
Corythalia dakryodes Bayer, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4806.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:722DB6C9-2C18-48EB-B202-7F2AFF47F49F |
|
persistent identifier |
https://treatment.plazi.org/id/03E68831-AF2C-4BE2-80CF-130F855A5819 |
|
taxon LSID |
lsid:zoobank.org:act:03E68831-AF2C-4BE2-80CF-130F855A5819 |
|
treatment provided by |
Plazi |
|
scientific name |
Corythalia dakryodes Bayer |
| status |
sp. nov. |
Corythalia dakryodes Bayer View in CoL , sp. nov.
Figs 1 View FIGURE 1 D–E, 28A–C, 62E, 73A–B, 77A
urn:lsid:zoobank.org:act:03E68831-AF2C-4BE2-80CF-130F855A5819
Type material. Holotype: ♀, COLOMBIA [most likely the locality within Colombia is the following: Departa- mento del Magdalena: Zona Bananera : S of Ciénaga, roughly 74°13’W, 10°59’N within an area of 30 km southextension and 5 km west- and 5 km east-extension (in the southern section 10 km east-extension), roughly 20–50 m a.s.l.]; collected in Hamburg Harbour (Germany) with banana transportation, G. Schmidt leg. 13 June 1952, G. Schmidt det. as C. dimidiata Simon , deposited 1972, SMF 25804 GoogleMaps . Paratype ♀ with the same data as holotype, SMF 25804 (put in separate vial in the course of the present study) GoogleMaps .
Additional material. COLOMBIA: Departamento del Magdalena: Santa Marta, Rodadero : 1 ♀, Borys Mal- kin leg. 23–31 May 1968, AMNH-IZC 00327104 . ECUADOR: Santo Domingo de los Tsáchilas: Santo Domingo de los Colorados , about 550 m, ca. 0°15’S, 79°10’W: 1 ♀, W. Weyrauch (with highest likelihood) leg., SMF (col- lection number not yet given) GoogleMaps .
Etymology. The specific name refers to the tear-like (drop-like) secondary spermathecae of the female holotype (Ancient Greek “dakryodes” means “like tears”, “like drops”); term in apposition.
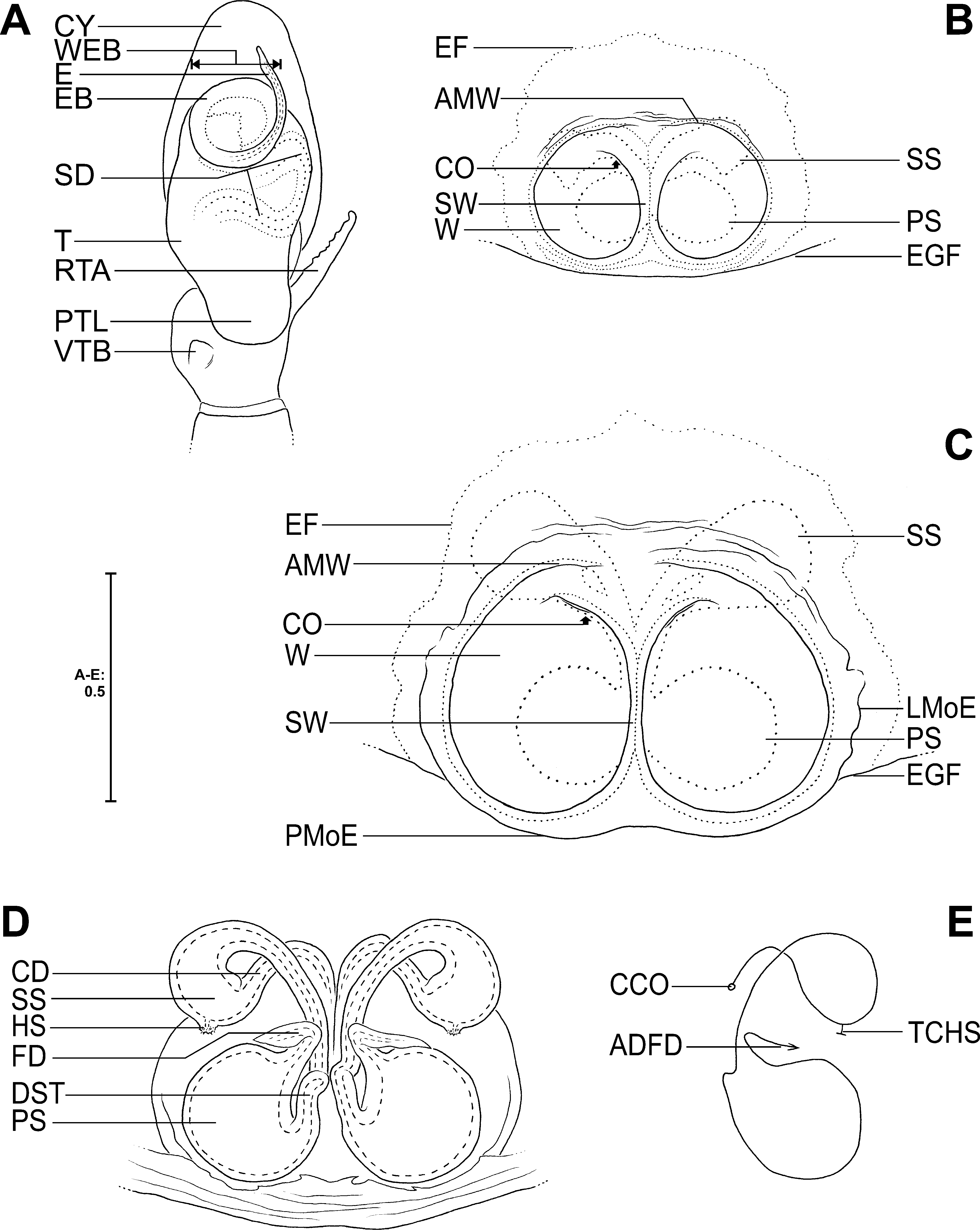
Diagnosis. Females distinguished from those of all other Corythalia species by the following characters in combination: epigynal windows (W) elongated oval; very small gap antero-laterally between lateral margin and anterior margin of W (the latter extremely diverging anteriorly and then running even postero-laterally) ( Figs 28A View FIGURE 28 , 73 View FIGURE 73 A–B); secondary spermathecae (SS) elongated drop-shaped ( Figs 1D View FIGURE 1 , 28B View FIGURE 28 , 77A View FIGURE 77 ); primary spermathecae (PS) as long as broad and not extending beyond posterior margins of epigynal windows; final section of copulatory duct (before meeting SS) running latero-posteriorly; connective ducts at final sections (before meeting PS) with loop-like windings ( Figs 1 View FIGURE 1 D–E, 28B–C, 77A).
Description. Male unknown.
Female (measurements of holotype first, those of paratype in parentheses): total length 8.3 (8.4), carapace length 3.9, maximal carapace width 3.1 (3.2), width of eye rectangle 2.3 (2.4), opisthosoma length 3.6, opisthosoma width 2.4 (2.5), fovea length 0.28 (0.33). EYES: AME 0.66 (0.70), ALE 0.39 (0.40), PME 0.14 (0.11), PLE 0.37 (0.38), AME–AME 0.07 (0.10), AME–ALE 0.12 (0.18), PME–PME 2.10 (2.21), PME–PLE 0.40 (0.44), ALE–PLE 1.05 (1.04), PLE–PLE 1.86 (1.97), clypeus height at AME 0.37 (0.36), clypeus height at ALE 0.88 (0.86). Cheliceral furrow with 1 promarginal and 1 retromarginal teeth. SPINATION: palp: no spines. Legs: femur I 1500, II 1600, III 1500 (1600), IV 0600 (1600); patella I 1000, II 1000 (1000{1010}), III–IV 1010; tibia I 2015, II 3025, III–IV 3133; metatarsus I–II 2024, III 3044 (3134), IV 5054 (4044). MEASUREMENT OF PALP AND LEGS: palp 3.3 (3.5) [1.4 (1.3), 0.5 (0.6), 0.4 (0.5), 1.0 (1.1)], I 6.2 (6.8) [2.1 (2.2), 1.2 (1.3), 1.3 (1.4), 1.0 (1.2), 0.6 (0.7)], II 6.3 (6.7) [2.1 (2.2), 1.3, 1.2 (1.3), 1.1 (1.2), 0.6 (0.7)], III 7.5 (8.1) [2.3 (2.6), 1.3 (1.4.), 1.4 (1.5), 1.6, 0.9 (1.0)], IV 7.9 (8.3) [2.5, 1.2, 1.6 (1.8), 1.7 (1.8), 0.9 (1.0)]. LEG FORMULA: 4321 (4312). COPULATORY ORGAN: epigyne with oval elongated epigynal windows (W) with very small gap antero-laterally between anterior and lateral margin of W ( Figs 28A View FIGURE 28 , 73 View FIGURE 73 A–B); W so to speak with double anterior margin; septum very narrow; epigynal field clearly broader than long, surrounding epigyne just narrowly ( Figs 28A View FIGURE 28 , 73 View FIGURE 73 A–B); structures of vulva visible through epigynal cuticle, primary spermathecae (PS) filling more than posterior half of W ( Figs 28A View FIGURE 28 , 73 View FIGURE 73 A–B). Vulva with large spherical PS; secondary spermathecae (SS) elongated tear-drop-shaped, with heads of spermathecae located posteriorly ( Figs 1 View FIGURE 1 D–E, 28B–C, 77A); connective ducts between both spermathecae running diagonally (with a moderate bent) from antero-lateral to postero-medial, with a small loop-like winding finally, before meeting PS (postero-) medially; copulatory duct with an arcuate course from copulatory opening antero-medially to SS ( Figs 1 View FIGURE 1 D–E, 28B–C, 77A); fertilisation ducts arising centro-anteriorly on primary spermathecae, initially running medio-anteriorly, then bent and directed transversal laterally ( Figs 1 View FIGURE 1 D–E, 28B–C, 77A). COLOURATION: see genus description for conservative aspects. Carapace red-brown ( Fig. 62E View FIGURE 62 ). Legs quite light yellowish brown to red-brown ( Fig. 62E View FIGURE 62 ). Opisthosoma like noted in genus description under general dorsal colouration, except for anterior transversal band missing, chevron-like patch in central band relatively light red-brown, but recognisable as such ( Fig. 62E View FIGURE 62 ).
Intraspecific variation of female copulatory organs. Female holotype ( Figs 28A View FIGURE 28 , 73A View FIGURE 73 ) with slightly longer epigynal windows than in paratype ( Fig. 73B View FIGURE 73 ); posterior half of epigyne in paratype slightly broader than anterior half ( Fig. 73B View FIGURE 73 ), in holotype anterior and posterior halves equal in width ( Figs 28A View FIGURE 28 , 73A View FIGURE 73 ). Primary spermathecae in holotype ( Figs 28 View FIGURE 28 A–B, 73A, 77A) in relation to the width of epigynal windows slightly larger than in paratype ( Figs 1D View FIGURE 1 , 73B View FIGURE 73 ). In paratype secondary spermathecae (SS) slightly less elongated and connective ducts subdistally with clearly less distinct loop ( Fig. 1D View FIGURE 1 ), whereas in holotype SS being more elongated and connective duct finally with conspicuous loop ( Figs 28B View FIGURE 28 , 77A View FIGURE 77 ).
Remarks. The two female types were collected by G. Schmidt 1952 at the harbour in Hamburg. Several years later G. Schmidt identified them as C. dimidiata Simon, 1901 . As Schmidt traced the origin of the charge (of banana fruits) as ‘Colombia’, he must have checked the catalogues on spider literature of species known at that time, i.e. Roewer (1954a, b) and Bonnet (1956). In these publications the land/island/region/continent of distribution is listed for each species. Schmidt might have found that C. dimidiata was the only species listed for Colombia except C. electa , which could be excluded from the further identification process because of the different dorsal colouration of the females. Note: at that time the females of C. electa were still misinterpreted; in the present study we found that the female paralectotypes were definitely misidentified, and, in fact, are C. spiralis (see under C. electa below). Corythalia dimidiata , however, is a nomen nudum, which is definitely not identifiable and recognisable (see under ‘ nomina nuda ’ below).
The locality in Colombia from where the type specimens originated is most likely within the Zona Bananera south of Ciénaga (see material list above) as by far the most Colombian bananas that are exported abroad are planted in that region.
As long as the conspecific male of C. dakryodes Bayer , sp. nov. remains undiscovered, it is difficult to predict possible relationships of this species. The species belonging to the C. waleckii species group ( C. waleckii , C. electa , C. tropica , C. fulgipedia , C. longiducta sp. nov. and others) might be closely related as they have the following characters in common: elongated drop- or kidney-shaped secondary spermathecae, quite long connective ducts, more or less round primary spermathecae and elongated epigynal windows. However, in this new species the distal sections of the copulatory ducts are running latero-posteriorly, because of the very far anterior situated copulatory openings. At least this aspect would rather argue against a close relationship with the species of the C. waleckii species group.
Distribution. Colombia, Ecuador.




| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |