
Pseudomaevia Rainbow, 1920
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3811.3.10 |
|
publication LSID |
lsid:zoobank.org:pub:09B993A9-EC44-4BAE-9B17-EB82470C0CA5 |
|
DOI |
https://doi.org/10.5281/zenodo.6135331 |
|
persistent identifier |
https://treatment.plazi.org/id/03D88787-D24C-7C55-FF5B-FB6F806FFECC |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudomaevia Rainbow, 1920 |
| status |
|
Pseudomaevia Rainbow, 1920 View in CoL
Type species: Pseudomaevia cognata Rainbow, 1920 by monotypy.
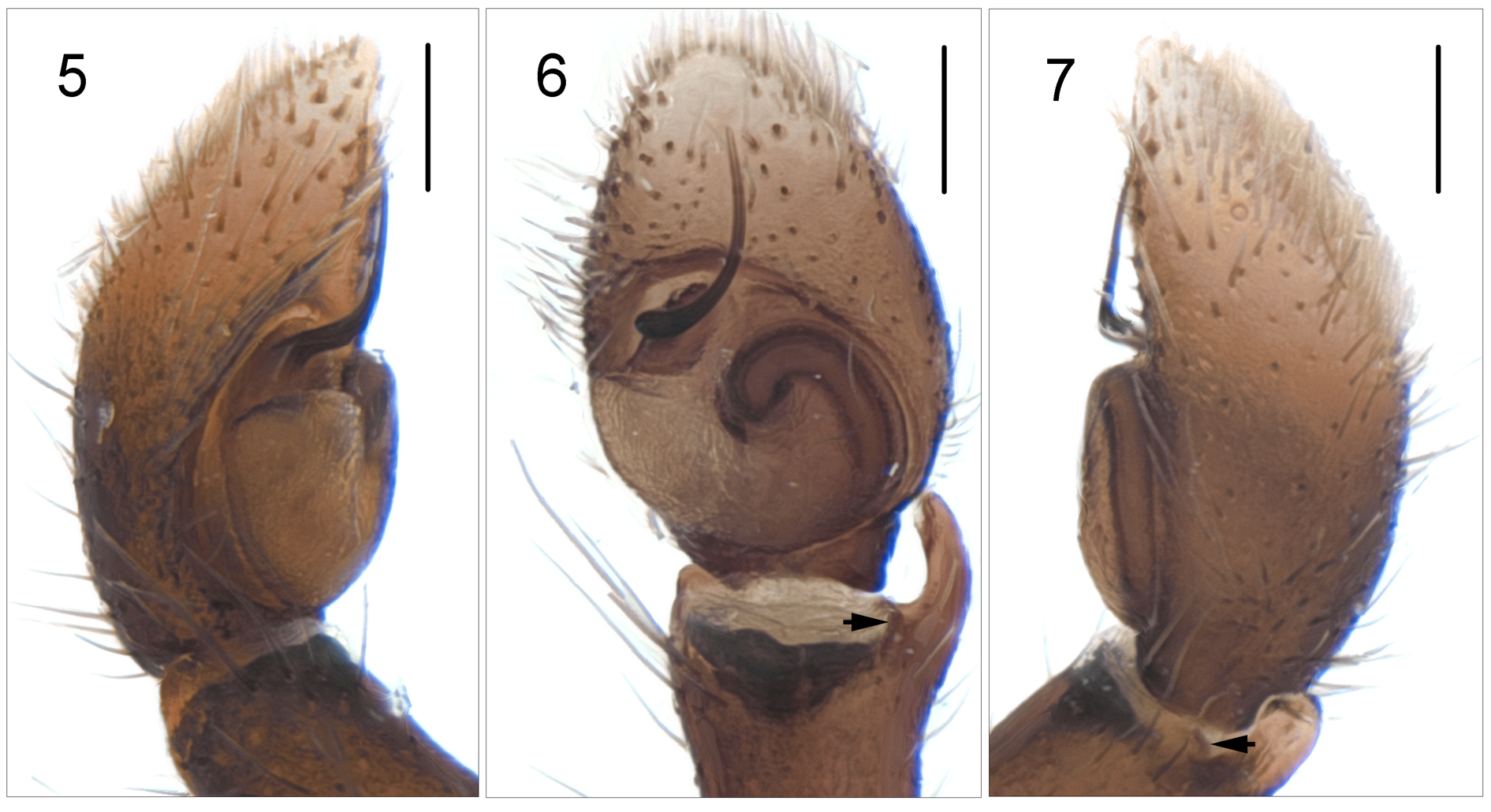
Remarks. As well as the male type specimen of P. cognata , a number of extra specimens collected more recently are available. These include females, which are described here. The sexes are considered conspecific because of the similar body forms and color patterns, plus the absence of congeneric species on the island. Female genitalic anatomy is unusual in that there is a well-developed diverticulum or gland arising from the insemination duct not far from the copulatory opening and the insemination duct joins the fertilization duct without any apparent spermatheca ( Figs 13–14 View FIGURES 12 – 15 ). The absence of a diverticulum and the presence of a well-developed spermatheca in P. insulana Berland, 1942 and, presumably, P. i. aorai Berland, 1942 , shows that they are not congeneric with P. cognata ( Żabka 1988) .
A possibly related genus is Corambis Simon, 1901 from New Caledonia and the Loyalty Islands ( Żabka 1988, Szűts 2002). Corambis and Pseudomaevia are similar in sharing an elongate general morphology with long strongly built L1’s and in the presence of poorly defined spermathecae with diverticulae and very long, laterally-directed, fertilization ducts. However the ‘flow’ through the systems is in opposite directions, with medially placed fertilization ducts and lateral insemination openings in Corambis , while it is the insemination openings that are medially placed in Pseudomaevia , with lateral fertilization ducts ( Figs 13–14 View FIGURES 12 – 15 ). In the male, in Corambis , unlike Pseudomaevia , fringing is present on L1 and there are four large spines on the tibia of L1. In Pseudomaevia ( Fig. 6 View FIGURES 5 – 7 ) there is a pair of tibial apophyses on each palp and the long embolus arises from a distinct mound behind the prolateral edge of the tegulum. This then moves away from the tegulum before forming an anti-clockwise quarter circle. There is no conductor separate from the embolus, and no proximal lobe on the tegulum in Pseudomaevia .
There are no other obviously related genera, most similar seemingly Baviola Simon, 1898 from the Indian Ocean, and Spilargis Simon, 1902 from Indonesia, New Guinea and the Pacific ( Prszyński 2014). However these are both considered members of the Euophryinae while Pseudomaevia is not: for example, the peripheral position of the sperm duct is unlike the distinctive meandering course followed by euophryines. Pseudomaevia also differs from these genera in the absence of obvious spermathecae in the female and the presence of a pair of, rather than a single, tibial apophyses in the male. Several Australian genera, namely Mopsoloides, Mopsus and Sandalodes also lack obvious spermathecae. However, in these genera the ducts have a convoluted structure with the copulatory openings near the posterior edge of the fossa, unlike Pseudomaevia . They are also unident and have a high rounded cephalothorax rather than fissident with a low, flat, parallel-sided cephalothorax.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.