
Setaria tundra, Isaichikov & Rajewskaja, 1928
|
publication ID |
https://doi.org/ 10.1016/j.ijppaw.2017.01.002 |
|
persistent identifier |
https://treatment.plazi.org/id/03D8879C-3171-FFD9-FCE1-F9D5FC20FA4E |
|
treatment provided by |
Felipe |
|
scientific name |
Setaria tundra |
| status |
|
3. Results View in CoL and discussion
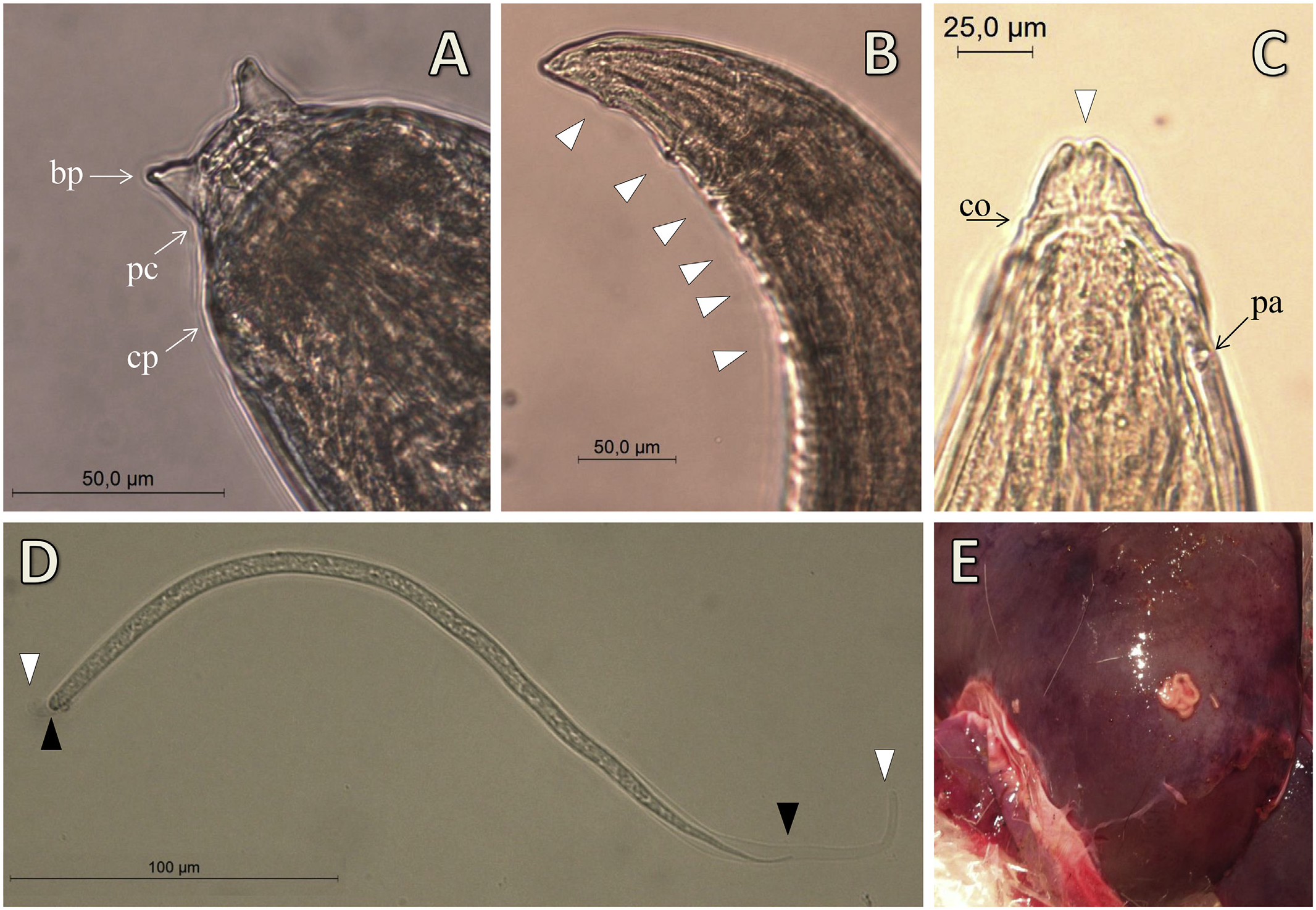
The morphology of worms isolated from the peritoneal cavity was identical to S. tundra by light microscopy ( Fig. 2 View Fig ). This was confirmed by sequences of the 12S rRNA and cox1 genes, which were 99.1 ‾ 99.8% identical to previously published S. tundra isolates from Germany, France, Italy, Spain and Finland. Phylogenetic trees constructed based on 12S rRNA and cox1 sequences were similar in topology and therefore only one tree is presented ( Fig. 3 View Fig ). Accordingly, the Danish isolates described here were clearly grouped with other isolates of S. tundra from Europe in one node that had high bootstrap values for NJ (0.99). However, sequence analysis showed slight variability within the Danish isolates, which was reflected in their topology in the phylogenetic tree. For example, the Danish isolates 2013-410-1 & 2, which geographically originated from Zealand (an island) were more closely related to the isolate 2010- 1233 that originated from Jutland (a peninsula) than the other isolate (2012-1665) from Zealand. The Danish isolate 2012-1665, with worms encapsulated in the liver capsule, had 0.5% variation in the cox1 compared to other Danish isolates, but had a lower level of variability (0.2 ‾ 0.3%) and was phylogenetically more closely related to other European isolates ( Fig. 3 View Fig ). Despite the low bootstrap values, the general topology of the Setaria spp. resembles a previously published tree by Alasaad et al. (2012).
This is the first report of S. tundra in Denmark. Worms of Setaria transcaucasica were earlier recovered from 41 out of 76 roe deer (53.9%) collected from the island LaesØ in Denmark ( Korsholm, 1988). In that study, the worms recovered from younger animals were encapsulated in different visceral organs and tissues, whereas in older animals the worms were found free in the peritoneal cavity. It is hard to determine if the proposed identification of the parasites was valid in the study by Korsholm (1988) because of the similar position of the worms in the subcapsular layer of visceral organs in that study in comparison to the current case 3 (Isolate 2012-1665) and scarce information can be found in the literature about the vectors, mode of transmission or morphology for S. transcaucasica . Indeed the taxonomic status of this parasite is highly ambiguous. Thus it is possible that these findings were actually S. tundra . Certainly, considering the presence of S. tundra to the north and east of the country it is unlikely that the parasites found in the current study represent very recent introductions into Denmark. Nonetheless, the existence of an isolated island with a remarkably high prevalence of a vector-borne filarial nematode may present a model for studying the transmission dynamics of that parasite.
Infections with S. tundra are of particular interest to game meat and fur retailers ( Rehbinder, 1990; Laaksonen et al., 2007). Reports from Finland linked poor body condition, under-developed fur in winter, dry fur, and reduced mean slaughter weight of reindeer infected with S. tundra ( Laaksonen et al., 2007) . In roe deer, most infections with S. tundra have less impact on meat and coat quality but if animals are severely afflicted approximately 152 metric tons of game meat ( DVFA, 2012), at an approximate value of 8,8 million Euros could be potentially affected. In Denmark, the annual hunting yield of roe deer is over 125,000 ( Asferg, 2012). Infected viscera from reindeer may be condemned at meat inspection, but the carcass is considered fit for human consumption even without heat treating ( Laaksonen et al., 2007). Based on the present results we consider the findings of S. tundra incidental and not associated with the emaciation, which was observed in three of the six cases (1, 2 & 6). In one animal (case 1) massive excretion of Giardia spp. cysts was seen and Cryptosporidium spp. oocysts were also detected. Another animal (case 6) was excreting high numbers of gastrointestinal nematodes. Only a single S. tundra was uncovered in the third animal (case 2) and no obvious parasitological explanation could be found for the observed enteritis and emaciation. Therefore the clinical signs may have been caused by other non-parasitic pathogens. Due to the low number of animals and the protracted study period nothing can be concluded regarding the general parasitological findings. It is worth noticing though that the zoonotic pathogens Cryptosporidium spp. and Giardia spp. ( García-Presedo et al., 2013) and/or I. ricinus , which is an important vector for several zoonotic agents ( Kauffmann et al., 2016; Scheid et al., 2016), were detected in all of the emaciated roe deer.
The spatio-temporal distribution and transmission of vector-borne diseases are highly affected by climatic change ( Hoberg et al., 2008). Previous outbreaks of setariosis in Scandinavia have been associated with unusually warm weather, and given the right circumstances the parasite has demonstrated its capacity to increase its geographic range considerably ( Rehbinder et al., 1975; Laaksonen et al., 2009). Additional mosquito species ( Coquillettidia richiardii and Ochlerotatus annulipes ) not previously known as S. tundra vectors have been observed to carry the parasite in Hungary, potentially increasing the vector species this parasite utilizes and it is likely that S. tundra is not vector specific ( Laaksonen et al., 2009; Angelone-Alasaad et al., 2016). This expected plasticity in vector species is concerning considering the likelihood that global warming will expand the range of various mosquito species into northern latitudes ( Dupouy-Camet, 2016).
While surveillance is based on voluntary submissions to the National Veterinary Laboratory nothing is known about prevalence of S. tundra in roe deer, fallow deer ( Dama dama ) and red deer ( Cervus elaphus ) (the three cervid species present in Denmark), although the distant geographical origin of the current cases in conjunction with their relatively diverse molecular characterization is further indicative of a well-established population. This does not preclude, however, increasing prevalence in the future as a response to climate change.
The distribution and abundance of hosts and vectors can explain the spatio-temporal presence of S. tundra . The outbreak of setariosis in the 1970s in Scandinavia was reported a few years after the introduction of roe deer into the same area ( Haugerud, 1989), which suggest a likely role of roe deer in the dissemination of this parasite ( Rehbinder et al., 1975; Laaksonen et al., 2007). The species is considered to be the predominant reservoir in Finland, and in Germany where prevalence ranges from 1.6% in North Rhine-Westphalia to up to 12.3% in northern Bavaria in roe deer ( Czajka et al., 2012). With a reported mean daily range of 8.5 ha in roe deer ( Jeppesen,1990) and an open land border between Jutland and Germany, frequent crossings are likely to occur and are an obvious possible route of the parasite into the country. How the parasite was introduced onto the island of Zealand is not clear but the heterogeneous genetic profiles found on the island could indicate multiple introductions. In northern Finland the increased prevalence of S. tundra in slaughtered reindeer ( Laaksonen et al., 2007) was linked to aggregation of reindeer in herds in mosquito-rich wetlands ( Laaksonen et al., 2009). The risk for transmission is highly enhanced when susceptible hosts are aggregated ( Opara and Fagbemi, 2008). Given that S. tundra is not vector specific, higher rate of transmission of this parasite is accordingly expected in woodlands that are close to water, where ungulates are aggregated in large numbers.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |