
Austrominius Buckeridge 1983
|
publication ID |
https://doi.org/ 10.5281/zenodo.275597 |
|
DOI |
https://doi.org/10.5281/zenodo.6196929 |
|
persistent identifier |
https://treatment.plazi.org/id/03D887BC-2162-F55E-FF60-B43EFB01FB4A |
|
treatment provided by |
Plazi |
|
scientific name |
Austrominius Buckeridge 1983 |
| status |
|
Genus Austrominius Buckeridge 1983
Elminius (Austrominius) Buckeridge, 1983a: 354 .
Diagnosis: Four-plated elminiines with thin-walled parietes; paries smooth internally, lacking “an inflected basal margin”; scutum without adductor ridge, articular ridge moderately prominent, adductor pits for depressor muscles absent; tergum with articular furrow wide, spur confluent with basi-rostral angle.
Distribution and age: Littoral, low energy environments. Recent: Southwest Pacific.
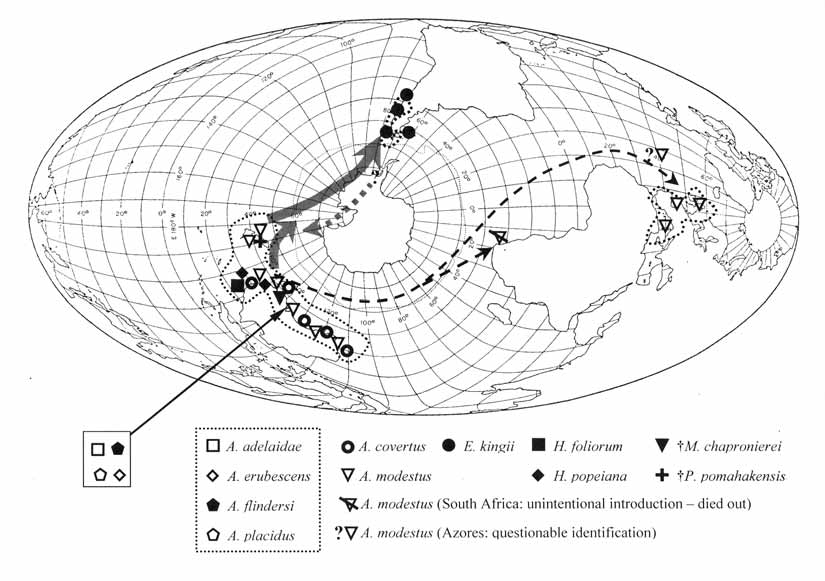
Type: Elminius modestus Darwin, 1854 . Recent: Australia, New Zealand [A common fouling species; introduced to southern England during the 1940s in war-time shipping convoys and more recently from Dunstaffnage, western Scotland (JB pers. obs., 1999)]. The distribution along the Atlantic seaboard is now fairly extensive, e.g. O’Riordan and Ramsay (1999), who recorded it from the Portuguese coast. They mention its occurrence in the Mediterranean (Zibrowius, pers. com.), but it is apparently known only from a very restricted distribution at Thau Lagoon, French Mediterranean (JB pers. obs., 1996, courtesy H. Zibrowius, and Zibrowius in Streftaris et al., 2005). The Thau population is small and may depend on yearly imports of oysters from the Atlantic coast of France (Zibrowius, pers. com). More recently, A. modestus has been report attached to drifting plastic as far north as the Shetland Islands ( Barnes and Miller, 2005).
Remarks: In the introduction we documented, as best we could, how the subgenus Austrominius became recognized at the full generic status, because it occurred more or less spontaneously rather than formally. This was done for historical rather than practical concerns, for Article 43.1 (Principle of Co-ordination) of the International Code of Zoological Nomenclature (1999) states that “a name established for a taxon at either rank in the genus group is deemed to have been simultaneously established by the same author for a nominal taxon at the other rank in the group; both nominal taxa have the same type species, whether it was fixed originally or subsequently.” Therefore, even though it is unnecessary, we consider it appropriate to formally endorse the full generic status of Austrominius , especially since it is being done in the same paper in which † Matellionius is being formally elevated.
Until Bayliss (1988, 1994), Austrominius had been a relatively small genus, but curiously his proposal of four new species did not attract attention at the time. His proposed species, Elminius adelaidae , E. eburescens , E. flindersi and E. placidus , all from the intertidal of South Australia, were then assignable to the subgenus Austrominius , which is now recognized as a full genus and, henceforth, they are so referred to herein ( Figures 1 View FIGURE 1 , 2; Table 2). Populations of all four of Bayliss’s taxa, along with Austrominius modestus , are known to occur within 50 km of each other ( Bayliss, 1994: Figure 3 View FIGURE 3 ). Bayliss (1994: 123) produced a key, in which differentiation of taxa is determined primarily on the basis of shell colour (although all of Bayliss’s taxa tend to be white or pale colours) and on pages 117 and 122, noted that it may be difficult to distinguish specimens that have worn or stained shells. Bayliss also used rather subtle variations in opercular valve morphology, minor variations in the colour patterns of the tergo-scutal flaps and slight differences in cirrial counts, but we fail to see how these could be considered specifically significant. Recent papers, such as Marchinko (2007), demonstrate that cirral counts in Balanus glandula Darwin, 1854 vary considerably in response to spatial and temporal differences in wave exposure; we suspect that ecotypic variation explains Bayliss’s forms, since they could be expected within a single species. There are numerous incidents in cirripedes where taxonomic splitting has been shown to have been premature: Van Syoc (1992) demonstrated that Amphibalanus saltonensis (Rodgers, 1949) (from the Salton Sea) was not a valid taxon, as specimens transferred to Mission Bay in California quickly established themselves as Amphibalanus amphitrite ( Darwin, 1854) ; Ross (1999) split Tetraclita rufotincta Pilsbry, 1916 into three species, which were shown in Appelbaum et al. (2002) to be only two species that nonetheless showed considerable “phenotypic plasticity”; thus we are not confident about Bayliss’ proposed species. Indeed, there is clearly a place for molecular techniques to clarify these taxa – and ideally this should be part of a genetic analysis of the Elminiinae as a whole. This will be the subject of a further paper2.
Hutton (1879) described two species of Austrominius from New Zealand: as Elminius sinuatus (from Wellington) and as Elminius rugosus (from The Bluff). Foster (1978) synonymised the former with E. modestus and confirmed the latter as a junior synonym of Epopella plicata . A further “species”, Austrominius australis , is listed in a table in Claessens (2005). This material was removed from the surface layers of a core sample collected in west Auckland, New Zealand and as the author provides no rationale for the name, it is here concluded that the use of “ australis ” is a mistake, as the material is almost certainly Austrominius modestus .
Late Eocene Miocene Holocene

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Cirripedia |
|
Order |
|
|
Family |
Austrominius Buckeridge 1983
| Buckeridge, John S. & Newman, William A. 2010 |
Elminius (Austrominius)
| Buckeridge 1983: 354 |