
Acorus calamus subsp. leaves
|
publication ID |
https://doi.org/10.1016/j.phytochem.2022.113318 |
|
persistent identifier |
https://treatment.plazi.org/id/03D987ED-FF94-AE58-FF99-F9CFBC7BF813 |
|
treatment provided by |
Felipe |
|
scientific name |
Acorus calamus subsp. leaves |
| status |
|
2.2. Volatile organic compound profiles in A. calamus leaves and rhizomes
Variation in the total content of A. calamus terpenes has been reported ( Raja et al., 2009; Dong et al., 2010; Parki et al., 2017). However, to the best of our knowledge, the role of TPSs in the formation of mono-, and sesquiterpene volatiles in A. calamus has not been reported.
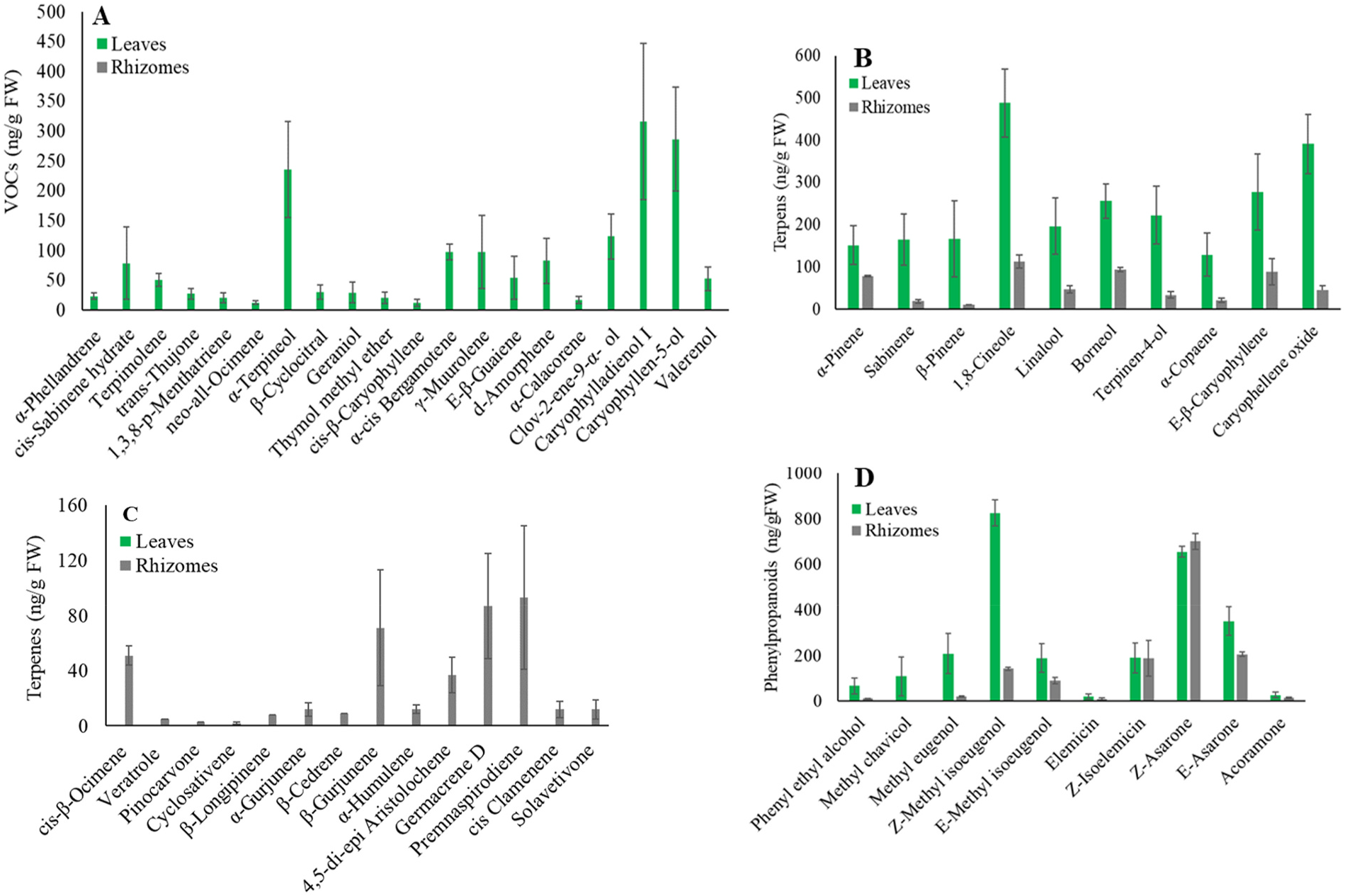
We examined the volatile compound composition of freshly harvested tissue from the leaves and rhizomes of A. calamus growing in our experimental station using SPME-GC-MS. A total of 110 volatile compounds were detected from the rhizomes and leaves of A. calamus , of which 32 were monoterpenes, 39 were sesquiterpenes, 17 were identified as phenylpropanoids, and 22 fatty acid-derived volatiles ( Fig. 1 View Fig and Supplemental Table 1 View Table 1 ). A. calamus leaves accumulated higher mono-and sesquiterpene levels than the rhizomes ( Fig. 1A View Fig and Supplemental Table 1 View Table 1 ). Moreover, significant differences in the composition of volatile compounds were observed between A. calamus leaves and rhizomes. For example, the most abundant leaf terpenes, caryophyllene oxide (3), Eβ- caryophyllene (4), caryophylladienol I (12), caryophyllen-5-ol (13), and α -terpineol (14) were absent or minor in A. calamus rhizomes ( Fig. 1A and B View Fig and Supplemental Table 1 View Table 1 ). The 1,8-cineole (15) ( 488 ng / g FW) was the most abundant terpene in the rhizome, while in leaves, it was 113 ng /g FW ( Fig. 1B and C View Fig and Supplemental Table 1 View Table 1 ). Compounds like cis-β -ocimene (16), β -gujeunene (17), germacrene D (18), and premnaspirodiene detected in rhizomes were absent in the leaves ( Fig. 1C View Fig and Supplemental Table 1 View Table 1 ). Phenylpropanoids showed significant differences between the A. calamus leaves and rhizomes ( Fig. 1D View Fig and Supplemental Table 1 View Table 1 ). For example, Z -methyl isoeugenol (19) was the most abundant volatile in the leaves, but was found at a much lower concentration in the rhizomes ( Fig. 1D View Fig and Supplemental Table 1 View Table 1 ). It was also noteworthy that the leaves accumulated a higher level of phenyl ethyl alcohol (20), methyl chavicol (21), E -methyl eugenol (6), and β -asarone (7) than the rhizomes. Additionally, A. calamus leaves and rhizomes accumulated similar levels of α -asarone (8) and Z -isoelemicin (22) ( Fig. 1D View Fig and Supplemental Table 1 View Table 1 ).
A. calamus View in CoL has three cytotypic forms distinguished by the chromosome number, and the triploid form is the most common and native to Asia ( Balakumbahan et al., 2010; Chandra and Prasad, 2017). Motley (1994) reported at least 185 VOCs from the essential oil of the triploid European A. calamus var. calamus View in CoL and tetraploid Indian A. calamus var. angustatus View in CoL . More than fifty VOCs were recorded from A. calamus View in CoL rhizomes ( Gyawali and Kim, 2009). β -Asarone (7) with 45% was the main compound in the A. calamus View in CoL rhizomes from Nepale ( Gyawali and Kim, 2009). β -Asarone (7) was the major compound of the essential oil of the triploid European A. calamus var. calamus View in CoL and tetraploid Indian A. calamus var. angustatus ( Motley, 1994) View in CoL . The content of β -asarone (7) and composition of VOCs in plant parts vary with the plants geographical conditions, plant age, climate, and ploidy ( Rajput et al., 2014). Furthermore, all angiosperm lineages have remnants of previous rounds of whole-genome duplication (WGD), emphasizing the importance of polyploidy as a driving force in plant evolution ( Leitch and Leitch, 2008). Because of the advantageous evolutionary success, polyploidy has multiple effects at different scales, from the molecular to the phenotypic level. Specifically, WGD and subsequent gene duplication are essential molecular evolutionary mechanisms underlying the wide range of plant VOCs ( Leitch and Leitch, 2008; Flagel and Wendel, 2009; Picazo-Aragon´es et al., 2020; da Silva et al., 2021; Madani et al., 2021).
The composition of VOCs shows a significant difference in various plant tissues, e.g., leaves, fruits, roots, and rhizomes of different species ( Chen et al., 2011; Falara et al., 2011; Koo and Gang, 2012; Yahyaa et al., 2015a,b; Yahyaa et al., 2015, 2015, 2016; Nawade et al., 2019). VOCs in the different tissues of A. calamus View in CoL might support multiple functions, and they can be involved in plant defense by directly repelling herbivores or pathogens or recruiting enemies of their aggressors to limit or eliminate further damage. It has been reported that the emission of terpene volatile compound mixtures from leaves, roots, and rhizomes tissues of many plant species serves as an essential ecological function ( Schnee et al., 2002; Rasmann et al., 2005).
Terpene oils produced in A. calamus have been traditionally used medicinally against different diseases, e.g., fever, asthma, bronchitis, cough, and digestive problems ( Rajput et al., 2014; Sharma et al., 2014; Chandra and Prasad, 2017; Khwairakpam et al., 2018), and they could also play interesting defensive roles in above- and below-ground biotic stress. Besides their ecological benefits, volatile terpene compounds are widely used by humans as flavors, fragrances, or pharmaceuticals ( Bohlmann and Keeling, 2008; Maffei et al., 2011; Nawade et al., 2020; Sharmeen et al., 2021).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |