
Heteromys nubicolens, Anderson & Timm, 2006
|
publication ID |
https://doi.org/ 10.1206/0003-0082(2006)3509[1:ANMSOS]2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.5649222 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87BD-FF85-256E-FCFD-FAD80026FD33 |
|
treatment provided by |
Felipe |
|
scientific name |
Heteromys nubicolens |
| status |
sp. nov. |
Heteromys nubicolens View in CoL , new species
Cloud-dwelling Spiny Pocket Mouse
Figures 3 View Fig , 5 View Fig , 6 View Fig , 7 View Fig
HOLOTYPE: KU 159025, nulliparous adult female; skin, skull, and postcranial skeleton in excellent condition; plus frozen tissues originally preserved in 95% ethanol (fig. 3). Collected on 16 October 2000 from COSTA RICA: PUNTARENAS: Monteverde, Monteverde Cloud Forest Reserve, Investigator’s Trail , 10 ° 189N, 84 ° 489W, at 1550 m elevation by Robert M. Timm and Christy M. McCain; original number RMT 4468.
PARATYPES: We designate as paratypes the following 24 specimens (adults in age classes 4–6; skins and skulls in good condition) from the Pacific slope or crest of the Cordillera de Tilarán in the Monteverde region, and housed in a variety of museum collections (appendix 1): COSTA RICA: PUNTARENAS: Monteverde , 1450 m, ROM 97307 ; Monteverde, Arthur Rockwell’s cafetales, 1400 m, UMMZ 115419 View Materials , 115420 View Materials ; Monteverde, Cerro Amigos , 1790 m, KU 142057 ; Monteverde, Hoge woods, 1420 m, LACM 64867 View Materials ; Monteverde, John Campbell’s woods, 1520–1580 m, FMNH 128417 View Materials , 128419 View Materials , 128420 View Materials , 128423 View Materials , 128425 View Materials ; KU 142791 ; LACM 64863 View Materials , 64865 View Materials ; MVZ 161224 View Materials , 161225 View Materials ; Monteverde, Monteverde Cloud Forest Reserve, Investigator’s Trail , 1550 m, KU 159022–159024 , 159026 , 159027 , 159029 ; MNCR 1336 View Materials ; Monteverde, Quebrada Quecha , KU 143337 ; Monteverde, Stella Wallace’s house, KU 143339 .
ETYMOLOGY: The adjective nubicolens , or ‘‘cloud-dwelling,’’ derives from the Latin nubes (cloud) and colo (dwell, inhabit) and is applied here in reference to the species’ distribution in cloud forests present on mountains that rise above the surrounding lowlands of northwestern Costa Rica ( Brown, 1956: 213, 478).
DISTRIBUTION: Known only from the Cordillera de Tilarán and Cordillera de Guanacaste of northwestern Costa Rica (fig. 4). In the Monteverde region of the Cordillera de Tilarán, it is found from 750 to 1840 m on the Caribbean slope and from 1350 to 1840 m on the Pacific slope. In the Cordillera de Guanacaste, the species has been collected at 1100–1500 m on Cerro Cacao and from 800 to 1200 m on Volcán Rincón de la Vieja–Volcán Santa María. See also Sympatry and Zones of Contact with Heteromys desmarestianus , below.
DIAGNOSIS: A species of spiny pocket mouse with adults showing the following combination of characters (figs. 3, 5, 6, 7): p4 (lower permanent premolar) with 3 lophs; P4 (permanent upper premolar) with long, curved fold in anterior border of posterior loph; mesopterygoid fossa V-shaped, with long, thin hamular processes of pterygoids; optic foramen small, with exterior margin formed by strong bar of bone; parietomastoid suture dipping well ventral to parietal crest posterior to its widest point; braincase not inflated; interorbit narrow; rostrum long and cylindrical; skull large (ONL 35.72–41.02 mm in adult specimens of age class 4; tables 1 View TABLE 1 , 2), elongated and relatively narrow; body size average to large for genus; dorsal pelage soft to moderately spiny; dorsal coloration dark brown and only faintly grizzled with thin ochraceous hairs intermixed among spines; no lateral ochraceous band present on flanks; plantar surface of hind feet naked.
DESCRIPTION: Dorsal pelage (fig. 5) soft to moderately spiny and dark brown (sharply contrasting with soft, pure white pelage of venter), only faintly grizzled with thin ochraceous hairs intermixed among spines (grizzling generally less pronounced along midline, producing slight dorsal stripe); ears dark brown to dark gray and small to medium in size; tail strongly bicolored for most of its length (then unicolored dark distally), generally longer than head-and-body length ( tables 1 View TABLE 1 , 2); patch of dark coloration present on dorsal and external surfaces of forearms, continuous with dark coloration of flanks; ventral and internal surfaces of forearms white; hind feet large (35 mm or greater in adults; tables 1 View TABLE 1 , 2), with naked plantar surface; skull (figs. 3, 6, 7) moderately large for genus ( tables 1 View TABLE 1 , 2); anterior half of premaxillary convex (inflated), forming a smooth (not stepped) lateral border of rostrum (in dorsal view); rostrum long and cylindrical (not tapered anteriorly or with dorsal flare anteriorly); nasals inflated anteriorly; interorbital constriction narrow; braincase narrow, not inflated; parietal and temporal crests weakly to moderately developed; interparietal moderately wide, often with slight anterior point; incisive foramina generally thin and slightly tapering anteriorly; no swelling at posteroventral border of infraorbital foramen 5; mesopterygoid fossa V-shaped, with long, thin hamular processes of pterygoids; shallow parapterygoid fossa; postalar fissure large and distinctly rounded anteriorly; optic foramen small, with exterior margin formed by strong bar of bone; parietomastoid suture dipping well ventral to parietal crest posterior to its widest point; anterodorsal lobe of periotic capsule of mastoid well developed, causing distinct undulation in posterior margin of squamosal (see schematic drawing in Anderson, 1999: 619); jugular foramen (anterolateral to occipital condyle) medium in size; dental formula: (incisors 1/1, canines 0/0, premolars 1/1; molars 3/3) × 2 5 total 20; molars moderately robust, with lophs of M2 subequal in width; lateral borders of lophs of molars smooth, not pointed; P4 with long, curved fold in anterior border of posterior loph, and with smoothly curved posterior border; p4 with 3 lophs; m3 not reduced, approximately same width as p4; angular process of mandible flat and blunt.
5 Erroneously listed as ‘‘anterodorsal border of infraorbital foramen’’ in Anderson (2003b). In most species of Liomys , a distinct swelling is present at the posteroventral border of the infraorbital foramen (generally anterodorsal to the incisive foramina). Such a swelling is never present in species of Heteromys . Other, more salient and consistent differences between the two genera are listed in Anderson (2003b: 11).
MEASUREMENTS OF THE HOLOTYPE (mm): total length 295, tail length 165, hind foot length 38, ear length 17, ONL 37.43, ZB 17.23, RL 17.45, NL 15.63, IOC 9.23, SB 16.12, MTR 5.38, IW 9.64, IL 5.26, PB 13.77, SD 11.42. Mass 68 g.
KARYOLOGY: Rogers (1989) reported the standard karyotype for two specimens now known as Heteromys nubicolens (from Provincia de Puntarenas: Monteverde, John Campbell’s woods, MVZ 161224 View Materials , 161225 View Materials ; appendix 1). This karyotype, the cytotype F of Rogers (1989), has a diploid number of 2n 5 60 and a fundamental number of FN 5 86 .
COMPARISONS: Externally, the dark brown, moderately grizzled pelage of Heteromys nubicolens differentiates it from most currently recognized species of the genus (fig. 5). Heteromys gaumeri , H. oasicus , and most populations of H. anomalus have pale brown dorsal pelage that is strongly grizzled with thin ochraceous hairs intermixed among the spines ( Anderson, 2003b). In addition, specimens of H. gaumeri possess a lateral ochraceous band that is absent in H. nubicolens . Heteromys nubicolens is much larger than the dwarf species H. oasicus (total length 227–250 mm for adults in age class 4; Anderson 2003b). Heteromys gaumeri is restricted geographically to Belize, Guatemala, and the Península de Yucatán in Mexico, and H. anomalus and H. oasicus inhabit only South America (Williams et al., 1993; Anderson, 2003b). Quite the reverse, H. australis , H. nelsoni , H. oresterus , and H. teleus show a dark slaty-gray, nearly uniform dorsal pelage, which separates them from the dark brown, moderately grizzled pelage of H. nubicolens . Heteromys teleus and H. australis are South American species, with the latter also showing a marginal distribution in extreme eastern Panama ( Anderson, 1999; Anderson and Jarrín-V., 2002). Both H. nelsoni and H. oresterus display extremely soft dorsal pelage and are distributed in restricted highland regions of extreme southeastern Mexico and southwestern Guatemala ( H. nelsoni ) and the Cordillera de Talamanca in Costa Rica ( H. oresterus ; Rogers and Rogers, 1992; Williams et al., 1993; Reid, 1997). However, H. nubicolens is quite similar in external morphology to species of the H. desmarestianus complex, which ranges from southern Mexico (Estado de Veracruz) to extreme northwestern Colombia (Williams et al., 1993). Many populations of this complex possess a lateral ochraceous band that is absent in H. nubicolens , but comparisons are best made using cranial characters.
Cranial characters serve to separate Heteromys nubicolens from all other species of the genus (figs. 3, 6, 7). Heteromys nubicolens differs from H. gaumeri by the narrow postalar fissure and strongly concave anterior termination of the premaxillary of H. gaumeri . The formation of the premaxillary in H. gaumeri creates a distinct step in the lateral border of the rostrum in dorsal view, a character lacking in the new species. Heteromys nubicolens contrasts with the South American species H. anomalus , H. australis , H. oasicus , and H. teleus by the especially small optic foramen and the straight, only moderately long fold in the anterior border of the posterior loph of P4 of those species. Heteromys nubicolens is average in size for the genus ( tables 1 View TABLE 1 , 2), not nearly as large as the massive H. nelsoni (Williams et al., 1993) .
Heteromys oresterus is similar to H. nubicolens in many respects but shows several clear differences. As mentioned above, H. oresterus has dark slaty-gray, nearly uniform dorsal pelage, in contrast to the dark brown, moderately grizzled pelage of H. nubicolens . Additionally, H. oresterus shows extremely soft dorsal pelage, whereas the dorsal pelage of H. nubicolens is soft-to-moderately spiny. Although spininess often varies clinally with elevation within a species of Heteromys (spinier in the lowlands and softer at higher elevations; Anderson, 1999: 619, 2003b: 23, 26; Anderson and Jarrín-V., 2002: 12), the difference in spininess between these two species is marked, despite the fact that they occur at broadly overlapping elevations. Cranially, H. oresterus differs by having an especially long, narrow rostrum (fig. 7) with a distinctive dorsal flare at the anterior termination of the rostrum (best observed in lateral view). Additionally, H. oresterus has deep parapterygoid fossae, in contrast to the shallow ones of H. nubicolens . In quantitative comparisons, H. oresterus averages larger than H. nubicolens in total length, head-and-body length, hind foot length, and occipitonasal length, but its average for the tail/head-andbody length ratio is smaller than that of H. nubicolens (P, 0.041 for significant comparisons, despite small sample size for H. oresterus ). Other quantitative comparisons between these two species were nonsignificant (P. 0.084; test not possible for mass; see tables 1–3 View TABLE 1 View TABLE 2 View TABLE 3 for descriptive statistics). In addition to these external, cranial, and mensural differences, H. oresterus and H. nubicolens also differ in standard karyotype; whereas the diploid number for each species is 2n 5 60, the fundamental number for H. oresterus is FN 5 78, in contrast to the FN 5 86 of H. nubicolens (Rogers, 1989) .
A member of the Heteromys desmarestianus complex comes into geographic contact with H. nubicolens in northwestern Costa Rica (see Sympatry and Zones of Contact with Heteromys desmarestianus , below) and merits close comparisons (figs. 6, 7). Currently, many species-level taxa are considered synonyms of H. desmarestianus (Williams et al., 1993; appendix 2), but none of them corresponds to the species here described as H. nubicolens . The two species present in northwestern Costa Rica are extremely similar externally, but differ in cranial size and proportions. Although both possess dark brown moderately grizzled dorsal pelage, that of H. nubicolens is generally less grizzled. In particular, very few ochraceous hairs are present along the midline of H. nubicolens , producing a gray dorsal stripe that is lacking in adjacent populations of H. desmarestianus . In addition, at Poco Sol (locality 7; appendix 1) near where the two species’ distributions apparently come into contact in the Peñas Blancas Valley (see locality 24 of H. nubicolens ), the ochraceous hairs of H. desmarestianus are especially pronounced around the neck, creating a ‘‘collar’’ not present in H. nubicolens (especially notable in the field; C.M. McCain, personal commun.). Otherwise, the two species are virtually identical in external coloration. Interestingly, H. nubicolens and H. desmarestianus exhibit vastly different behaviors when handled. Heteromys desmarestianus is quite vocal, uttering an array of audible sounds and actively struggling when held, whereas individuals of H. nubicolens make no audible vocalizations when handled and are quite passive (M. Klaus, personal commun.; C.M. McCain, personal commun.).
Quantitative comparisons document differences in size and shape between Heteromys nubicolens and H. desmarestianus . Although the two species overlap for mass and all external and cranial measurements, means were significantly different for each variable except ear length (P, 0.002 for significant comparisons; P 5 0.75 for ear length; table 1 View TABLE 1 ). Generally, H. nubicolens is larger, but least interorbital constriction, interparietal length, and parietal breadth average greater for H. desmarestianus (see also figs. 6, 7). Accordingly, although overlap occurs, the species also differ in mean for the ratios of least interorbital constriction/ occipitonasal length and parietal breadth/ occipitonasal length (P, 0.0001). Indeed, the species barely overlap in a plot of least interorbital constriction versus occipitonasal length (fig. 8). In addition, H. nubicolens shows a larger tail/head-and-body length ratio (P 5 0.0006).
Multivariate analyses also support the morphological distinctiveness of Heteromys nubicolens from H. desmarestianus . Only moderate overlap occurs between specimens of the two species in a plot of scores on the first two principal components of the PCA undertaken on specimens in age class 4 (fig. 9). The coefficients and loadings of PC1 are all of the same sign ( table 4 View TABLE 4 ). Hence, we interpret that PC1 represents a size axis (with larger specimens having lower scores, because the signs of the coefficients are all negative). In contrast, PC2 represents a shape axis (because some coefficients are positive and others negative; table 4 View TABLE 4 ).
In sum, adults of Heteromys nubicolens show larger overall body size than H. desmarestianus and differ in cranial proportions (figs. 6, 7). Although most measurements are larger for H. nubicolens , specimens of H. desmarestianus have wider interorbits, longer interparietals, and wider braincases (see parietal breadth; tables 1 View TABLE 1 , 2). In addition to these characters, direct visual comparisons of skulls indicate several further differences between the two species. The nasals flare laterally as they approach their anterior termination in H. nubicolens , whereas they are subparallel in H. desmarestianus . This character, combined with the overall greater development of the rostrum in H. nubicolens , yields a more massive rostrum that is both absolutely and relatively longer and less tapered anteriorly (in dorsal view) than that of H. desmarestianus . Because of its long rostrum, the interorbital constriction of H. nubicolens occupies a position proportionately more posterior than in H. desmarestianus . Additionally, associated with the wide interorbit and braincase of H. desmarestianus , the parietal and temporal crests tend to converge anteriorly in a V-shape in that species, in contrast to the generally hourglass-shaped crests of H. nubicolens . Overall, the parietal and temporal crests are more strongly developed in H. desmarestianus , forming a conspicuous shelf along the lateral borders of the interorbital constriction (whereas the corresponding crest is only weakly developed in H. nubicolens ). Ventrally, the parapterygoid fossa is shallow in H. nubicolens , but deep in H. desmarestianus . Finally, the basisphenoid and basioccipital are wider at the level of the basisphenoid–basioccipital suture in H. nubicolens than in H. desmarestianus .
GEOGRAPHIC VARIATION: Moderate geographic variation exists between samples of Heteromys nubicolens from the Cordillera de Tilarán versus those of the Cordillera de Guanacaste ( table 2 View TABLE 2 ). Means for a few measurements were significantly different in univariate comparisons of specimens from Monteverde (localities 30–32; Cordillera de Tilarán) with those from Cerro Cacao (locality 28; Cordillera de Guanacaste). Externally, only head-and-body length (P 5 0.01; longer at Cerro Cacao) and mass (P, 0.0001; larger at Cerro Cacao) differed. In two cranial dimensions reflecting aspects of the width of the neurocranium, the Monteverde sample averaged larger (squamosal breadth, P 5 0.027; interparietal width, P 5 0.0009). Finally, the samples differed in mean for maxillary toothrow length (P 5 0.0006; longer at Monteverde).
SYMPATRY AND ZONES OF CONTACT WITH HETEROMYS DESMARESTIANUS : Presently available specimens document three regions where the distribution of Heteromys nubicolens comes into geographic contact with that of H. desmarestianus , including one site of sympatry (fig. 4). The two species show parapatric distributions in the foothills of the Peñas Blancas Valley (on the Caribbean slope of the Cordillera de Tilarán in the Monteverde region) and at intermediate elevations on the slopes of both Cerro Cacao and Volcán Rincón de la Vieja–Volcán Santa María (in the Cordillera de Guanacaste; see also fig. 6). Similar parapatric distributions have been observed for other pairs of species in the subfamily ( Genoways, 1973; Rogers and Engstrom, 1992; Anderson, 1999; Anderson et al., 2002b; Anderson and Jarrín-V., 2002; Anderson and Martínez-Meyer, 2004).
In the Peñas Blancas Valley on the Caribbean slope of the Cordillera de Tilarán in the Monteverde region, the species’ distributions probably meet at ca. 750–1000 m (fig. 4). Along the main course of the Río Peñas Blancas, a recent standardized survey captured Heteromys nubicolens from the continental divide down to 750 m in elevation (localities 24–26, 30, 32; appendix 1; McCain, 2004, in press). In contrast, the same survey only encountered H. desmarestianus from 750 to 1050 m in elevation; these sites near the Poco Sol Field Station correspond to an area farther from the main valley (voucher specimens were collected at 750 m, locality 7, whereas captures of this species at 1000– 1050 m were released; McCain, 2004; C.M. McCain, personal commun.).
On the slopes of Cerro Cacao in the Cordillera de Guanacaste, the species turn over at ca. 900–1100 m (fig. 4). At Cerro Cacao (locality 28; appendix 1), the new species has been collected from 1100 to 1500 m. Heteromys desmarestianus is known from Maritza, Sendero Cacao at 600–900 m (locality 12) on the western slopes of Volcán Orosí and Cerro Cacao (which share a common base, with their peaks being connected by a broad connection at ca. 1000 m). This sendero (trail) runs from the Maritza field station south to the Cerro Cacao field station (Estación Mengo), following a gradual ascent to ca. 900 m before rising steeply to the Cerro Cacao field station. Elsewhere in the general vicinity of Cerro Cacao, H. desmarestianus also is known from 600 to 900 m at Pitilla, Sendero Carica and Pitilla, Sendero Orosilito (locality 11). These trails rise from the Pitilla field station, which lies on the northeastern slope of Volcán Orosı´.
On the slopes of Volcán Rincón de la Vieja– Volcán Santa María, the species’ distributions turn over at ca. 800–1100 m, and one site of sympatry is known (figs. 4, 6). Here, collections of Heteromys exist from two areas, the southeastern and southwestern versants of the mountain. On a southeastern ridge, H. nubicolens has been collected at 1100 m (locality 27; appendix 1). Just below that site, H. desmarestianus is known from the Quebrada Provisión, 900–1000 m (locality 5) and from the Hacienda Santa María (locality 14). Goodwin (1946) gave the elevation of Hacienda Santa María as 3200 ft (5 975 m; Harris [1943] reported these specimens but did not give an elevation). Hence, in this area, H. desmarestianus appears to replace H. nubicolens at 1000–1100 m.
Both species also are known from the southwestern slopes of Volcán Rincón de la Vieja–Volcán Santa María, including a site of sympatry at 800 m (note that locality 13 for Heteromys desmarestianus 5 locality 29 of H. nubicolens ; figs. 4, 6; appendix 1). In this area, H. nubicolens has been collected at both Pailas, Sendero Cráter, 1200 m and Pailas, Sendero Pailas, near Río Colorado, 800 m (locality 29). These two trails emanate from the Pailas field station. Heteromys desmarestianus is known from Pailas, Sendero Pailas, 800 m (locality 13). In addition, two specimens that we tentatively ascribe to H. desmarestianus were collected from Pailas, Sendero Pailas, near Río Colorado, 800 m (locality 13). The skulls of these two individuals (KU 158614, 158615; adult females in age class 5) are smaller than those of H. nubicolens in the equivalent age class. Furthermore, they display most cranial characters typical of H. desmarestianus . However, the parietal and temporal crests are hourglassshaped, and the rostrum is massive and not tapered anteriorly. We provisionally identify these specimens as H. desmarestianus , but raise the possibility that they may represent rare hybrids, present along a very narrow contact zone between the two species. Genetic studies are necessary to examine this possibility further.
NATURAL HISTORY: Whereas copious ecological information is available for Heteromys desmarestianus at La Selva (localities 18, 19; appendix 1) in Costa Rica ( Fleming, 1974a, 1974b, 1977, 1983; Fleming and Brown, 1975; Vandermeer, 1979; Vandermeer et al., 1979; Timm, 1994) and for a few populations in other parts of the range of this species complex (e.g., Fleming, 1970; Quintero and Sánchez-Cordero, 1989; Sánchez-Cordero, 1993; Sánchez-Cordero and Fleming, 1993; Brewer and Rejmánek, 1999; Brewer, 2001), relatively little has been published regardinthe species now known as H. nubicolens . Most of what is known derives from Monteverde and adjacent localities in the Cordillera de Tilarán. Heteromys nubicolens represents a common forest-floor species in the small mammal community at Monteverde ( Ashe and Timm, 1987a; Timm and LaVal, 2000a). At Monteverde, adult H. nubicolens have a home range of ca. 0.15 ha and do not shift their home ranges appreciably even over several months; immatures have a smaller home range of ca. 0.10 ha ( Anderson, 1982: 166). Breeding can occur during any month ( Anderson, 1982). This species was captured at all elevations in a transect from 1840 m in the Monteverde region down the Peñas Blancas Valley to 750 m on the Caribbean slope of the Cordillera de Tilarán ( McCain, 2004, in press).
Heteromys nubicolens appears to have a moderate tolerance for human-altered habitats. In many years of trapping at Monteverde by R.M.T., this species was often captured in primary forests above ca. 1500–1550 m, as well as in secondary forests 20–35 years old at ca. 1350–1550 m. However, despite extensive trapping in highly disturbed habitats, R.M.T. never captured H. nubicolens in pastures, farmed areas, or other highly disturbed areas. Some of the secondary forests at Monteverde represent former pastures that were planted with native tree species through projects aimed at restoring wildlife habitat or conserving soil and water. Other pastures were simply abandoned and allowed to regenerate on their own. Although Timm (1994: 232) reported ‘‘tremendous population fluctuations of spiny pocket mice at Monteverde’’ (see also Anderson, 1982), the species is generally captured in higher numbers in secondary forests than in primary ones (R.M.T., personal obs.).
Several decades of theriological research in the Monteverde area allow for characterization of the communities of small, nonvolant mammals in which Heteromys nubicolens occurs (Timm and LaVal, 2000a, 2000b; Woodman, 2000; Durden and Timm, 2001; Timm and Vriesendorp, 2003). In cloud forest, lower montane wet forest, and lower montane rainforest at Monteverde (ca. 1500–1850 m; Haber, 2000; Nadkarni and Wheelwright, 2000a) in the general vicinity of where the holotype of H. nubicolens was obtained, the following small, nonvolant mammals occur: the marsupials Caluromys derbianus , Didelphis marsupialis , and Marmosa mexicana ; the shrews Cryptotis nigrescens and C. merriami ; the squirrels Microsciurus alfari and Sciurus granatensis ; the pocket gopher Orthogeomys cherriei ; and the sigmodontine rodents Nyctomys sumichrasti , Oligoryzomys vegetus , Oryzomys devius (5 O. albigularis ), Peromyscus nudipes , Reithrodontomys creper , R. gracilis , Reithrodontomys sp. , Rheomys raptor , Scotinomys teguina , and Tylomys watsoni . Additional species of mammals that occur sympatrically with H. nubicolens in other (generally lower) parts of the Monteverde/Peñas Blancas region include: the marsupials Micoureus alstoni and Philander opossum ; the squirrel Sciurus variegatoides ; and the sigmodontine rodents Melanomys caliginosus , Oligoryzomys fulvescens , Oryzomys alfari , and O. bolivaris . In R.M.T.’s trapping at Monteverde, P. nudipes was usually the most common small rodent in habitats where H. nubicolens was found; Oryzomys devius and Scotinomys teguina were typically collected along with this species of Heteromys as well (see also McCain, 2004).
In contrast to the information available for the Cordillera de Tilarán, little has been published on the mammals of the Cordillera de Guanacaste and surrounding lowlands; hence, even basic natural history information is lacking for Heteromys nubicolens there. Studies of communities of small nonvolant mammals have been conducted recently by Marion Klaus (Sheridan College, Sheridan, WY) at various localities in the Área de Conservación Guanacaste (including Cerro Cacao and Volcán Rincón de la Vieja– Volcán Santa María). Similarly, Neal Woodman (Patuxent Wildlife Research Center, United States Geological Survey) conducted a series of mammalian inventories in the region in 1987–1988. Voucher specimens from those projects were included in the present taxonomic study; future work by Klaus and Woodman may provide ecological and natural history information on Heteromys nubicolens in this region.
ECTOPARASITES AND OTHER ASSOCIATED INVERTEBRATES: Two species of ectoparasites have been collected on Heteromys nubicolens . Both are ticks (Acarina: Ixodidae ): Ixodes venezuelensis (from KU 158611) and Dermacentor sp. (from KU 158610 and KU 159026). Notably, two species of amblyopinine beetles ( Coleoptera : Staphylinidae ) that are commensal with other small mammals ( Oryzomys devius , Peromyscus nudipes , and Reithrodontomys creper , and rarely on Tylomys watsoni ) at Monteverde have not been found on H. nubicolens ( Ashe and Timm, 1987a, 1987b, 1995; Timm and Ashe, 1988). Similarly, a tineid moth, Amydria selvae , is phoretic (a nonparasitic association where one organism attaches to the other for dispersal) on H. desmarestianus at La Selva ( Davis et al., 1986) but has not been found on Heteromys nubicolens at Monteverde (R.M.T., personal obs.; Timm et al., 1989: 116). Finally, whereas pseudoscorpions of the genus Epichernes have been reported as phoretic on H. gaumeri and Liomys salvini (the latter in northwestern Costa Rica; Muchmore, 1992), none has been found on H. nubicolens .
CONSERVATION STATUS: Although we do not consider that Heteromys nubicolens currently needs additional special protection, future studies of its distribution and population sizes will allow for evaluation based on IUCN (2001) criteria. 6 All data available from field collectors indicate that H. nubicolens inhabits only closed-canopy forests (similar to H. australis and H. teleus ; Anderson and Jarrín-V., 2002; Anderson and Martínez- Meyer, 2004), including both primary forests and secondary stands more than 20 years in age (see Natural History). Although middle- and high-elevation forests have been less affected by deforestation than lowland forests in Costa Rica, high average annual rates of forest conversion continue in the country (Sader and Joyce, 1988; Sánchez-Azofeifa et al., 2001). Fortunately, many areas in the range of H. nubicolens are protected by extensive private and governmental reserves. For purposes related to conservation, Costa Rica is divided administratively into a series of official conservation areas that cut across provincial borders ( Janzen, 2000; Savage, 2002: 53; Mata and Echeverría, 2004). Within each conservation area, some regions correspond to national parks or other governmentally protected areas, but other land remains in private hands (including private conservation reserves). Known localities of H. nubicolens fall in a complex of private reserves in the Monteverde region (in the Área de Conservación Arenal; Burlingame, 2000), as well as in the Parque Nacional Guanacaste and Parque Nacional Rincón de la Vieja (in the Área de Conservación Guanacaste; Janzen, 2000; Savage, 2002; Mata and Echeverría, 2004). In addition, yet-undiscovered populations of the species likely exist (see Biogeography, below) in the Parque Nacional Volcán Arenal, Parque Nacional Volcán Tenorio, and other governmentally protected areas in the Área de Conservación Arenal.



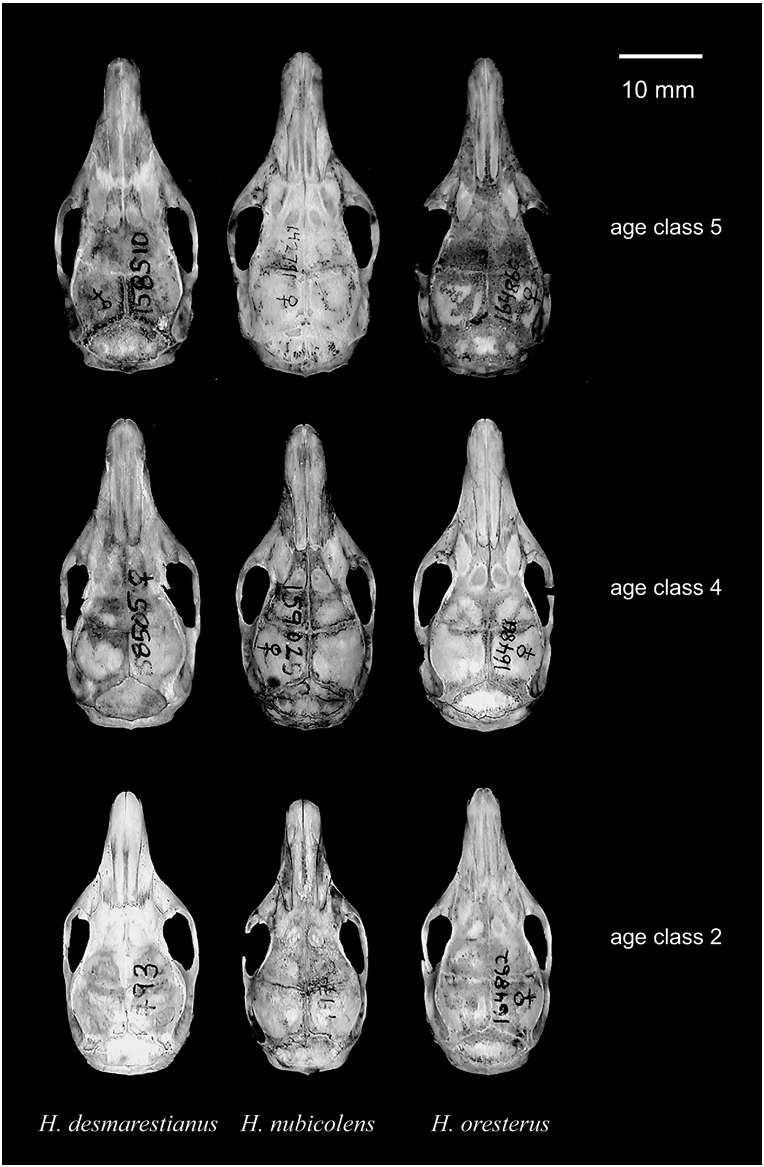
TABLE 1 Descriptive Statistics and Results of t-Tests Comparing Adults in Age Class 4 of Heteromys desmarestianus and H. nubicolens in Northwestern Costa Rica for External Measurements, Mass, Cranial Measurements, and Three Derived Ratiosa
| H. desmarestianus | H. nubicolens | |
|---|---|---|
| Total length**** | 292.2 ± 3.9, 46 (238–353) | 322.2 ± 2.7, 41 (291–358) |
| Head-and-body length** | 138.7 ± 1.9, 52 (96–169) | 146.4 ± 1.5, 52 (104–183) |
| Tail length**** | 152.8 ± 2.6, 46 (128–210) | 174.8 ± 1.7, 41 (150–197) |
| Tail/head-and-body ratio, × 100 (%; ***) | 110.0 ± 1.8, 46 (90.8–146.9) | 119.3 ± 1.9, 41 (95.6–179.8) |
| Hind foot length**** | 35.8 ± 0.3, 52 (32–40) | 38.7 ± 0.2, 51 (35–42) |
| Ear length (n.s.) | 17.9 ± 0.2, 51 (13–20) | 17.8 ± 0.2, 51 (15–21) |
| Mass (grams)** | 75.4 ± 2.7, 42 (37–110) | 86.6 ± 2.4, 50 (60–136) |
| Occipitonasal length**** | 36.506 ± 0.109, 176 (32.71–40.09) | 38.630 ± 0.177, 51 (35.72–41.02) |
| Zygomatic breadth**** | 16.519 ± 0.054, 134 (15.20–18.52) | 17.533 ± 0.085, 42 (16.40–18.51) |
| Rostral length**** | 16.558 ± 0.062, 176 (14.51–18.67) | 17.950 ± 0.110, 50 (15.82–19.36) |
| Nasal length**** | 15.283 ± 0.065, 175 (13.05–17.25) | 16.079 ± 0.112, 51 (13.89–17.78) |
| Least interorbital constriction**** | 9.789 ± 0.036, 176 (8.39–11.11) | 9.250 ± 0.067, 51 (8.20–10.40) |
| Squamosal breadth**** | 15.471 ± 0.032, 176 (14.40–16.59) | 15.932 ± 0.059, 51 (15.19–17.74) |
| Maxillary toothrow length**** | 5.252 ± 0.017, 176 (4.63–5.99) | 5.461 ± 0.035, 51 (4.97–5.92) |
| Interparietal width**** | 9.289 ± 0.042, 176 (8.02–10.75) | 9.715 ± 0.066, 51 (8.24–10.84) |
| Interparietal length**** | 5.288 ± 0.033, 176 (4.23–6.49) | 4.912 ± 0.071, 51 (3.82–6.11) |
| Parietal breadth**** | 14.286 ± 0.033, 176 (13.11–15.50) | 13.795 ± 0.045, 51 (12.85–14.60) |
| Skull depth**** | 11.202 ± 0.023, 176 (10.43–11.99) | 11.640 ± 0.032, 51 (11.00–12.09) |
| Least interorbital constriction/ occipitonasal length ratio, × 100 (%; ****) Parietal breadth/ occipitonasal length ratio, × 100 (%; ****) | 26.824 ± 0.076, 176 (23.93–28.79) 39.164 ± 0.093, 176 (35.38–41.80) | 23.944 ± 0.133, 51 (22.20–26.46) 35.742 ± 0.175, 51 (32.63–38.94) |
a Mean ± standard error, sample size, and observed range (minimum–maximum) are given for each species. Measurements with significant differences in two-tailed t -tests comparing means of the species are indicated (* 5 P # 0.05; ** 5 P # 0.01; *** 5 P # 0.001; **** 5 P # 0.0001; n.s. 5 P. 0.05). Note that whereas averages of most measurements are larger for H. nubicolens , averages for least interorbital constriction, interparietal length, and parietal breadth are larger in H. desmarestianus . Units are millimeters unless otherwise noted.
TABLE 4 Results of Principal Components Analysis for Adult Specimens in Age Class 4 of Heteromys desmarestianus and H. nubicolens from Northwestern Costa Ricaa
| Unit eigenvector | Loadings | |||||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| ln-Occipitonasal length | 20.364 | 0.096 | 0.025 | 20.941 | 0.201 | 0.027 |
| ln-Zygomatic breadth | 20.301 | 0.135 | 20.093 | 20.794 | 0.289 | 20.122 |
| ln-Rostral length | 20.466 | 0.175 | 0.066 | 20.906 | 0.277 | 0.057 |
| ln-Nasal length | 20.469 | 0.095 | 0.187 | 20.902 | 0.146 | 0.171 |
| ln-Least interorbital constriction | 20.215 | 20.209 | 0.652 | 20.440 | 20.339 | 0.654 |
| ln-Squamosal breadth | 20.187 | 0.047 | 20.046 | 20.755 | 0.151 | 20.095 |
| ln-Maxillary toothrow length | 20.268 | 0.112 | 20.108 | 20.669 | 0.229 | 20.138 |
| ln-Interparietal width | 20.303 | 20.007 | 20.638 | 20.564 | 20.009 | 20.583 |
| ln-Interparietal length | 20.215 | 20.919 | 20.179 | 20.288 | 20.934 | 20.118 |
| ln-Parietal breadth | 20.097 | 20.144 | 0.270 | 20.343 | 20.401 | 0.464 |
| ln-Skull depth | 20.198 | 0.090 | 0.015 | 20.772 | 0.283 | 0.027 |
| Eigenvalue (× 100) | 1.393 | 0.885 | 0.339 | |||
| % variation explained | 42.9 | 27.2 | 10.5 | |||
a Elements of the unit eigenvector and loadings are presented for the first three principal components. Cranial measurements were transformed to their natural logarithms (ln) before components were extracted from the covariance matrix. Eigenvalues (multiplied by 100) and the percentage of the variation among specimens explained are given for each axis.
TABLE 2 Descriptive Statistics for Adults in Age Class 4 of Heteromys desmarestianus from La Selva and H. nubicolens from Monteverde and Cerro Cacao in Northwestern Costa Ricaa
| La Selva | Monteverde | Cerro Cacao | |
|---|---|---|---|
| Total length | 265, 1 | 318.9 ± 4.1, 17 | 329.3 ± 3.5, 15 |
| (265) | (291–347) | (300–351) | |
| Head-and-body length | 131, 1 | 143.2 ± 2.2, 27 | 150.3 ± 1.5, 15 |
| (131) | (104–163) | (140–158) | |
| Tail length | 134, 1 | 175.1 ± 2.6, 17 | 179.1 ± 2.7, 15 |
| (134) | (153–192) | (155–197) | |
| Tail/head-and-body ratio, × 100 (%) | 102.3, 1 | 122.8 ± 3.9, 17 | 119.2 ± 1.8, 15 |
| (102.3) | (105.1–179.8) | (106.9–131.4) | |
| Hind foot length | 35, 1 | 38.9 ± 0.3, 26 | 38.7 ± 0.4, 15 |
| (35) | (35–41) | (36–42) | |
| Ear length | 17, 1 | 17.8 ± 0.2, 27 | 18.3 ± 0.3, 14 |
| (17) | (16–21) | (17–20) | |
| Mass (grams) | 62, 1 | 79.4 ± 2.2, 26 | 104.9 ± 3.5, 14 |
| (62) | (61–109) | (90–136) | |
| Occipitonasal length | 36.512 ± 0.123, 121 | 38.346 ± 0.210, 28 | 39.183 ± 0.376, 13 |
| (32.71–39.74) | (36.48–40.49) | (35.72–41.02) | |
| Zygomatic breadth | 16.359 ± 0.059, 85 | 17.413 ± 0.110, 24 | 17.831 ± 0.227, 8 |
| (15.20–17.83) | (16.40–18.31) | (16.52–18.51) | |
| Rostral length | 16.590 ± 0.069, 121 | 17.786 ± 0.125, 27 | 18.270 ± 0.234, 13 |
| (14.51–18.45) | (16.61–19.29) | (15.82–19.11) | |
| Nasal length | 15.365 ± 0.070, 121 | 15.999 ± 0.154, 28 | 16.265 ± 0.213, 13 |
| (13.21–17.25) | (13.89–17.78) | (14.25–17.41) | |
| Least interorbital constriction | 9.919 ± 0.038, 121 | 9.222 ± 0.088, 28 | 9.419 ± 0.093, 13 |
| (8.75–11.11) | (8.37–10.19) | (8.78–10.04) | |
| Squamosal breadth | 15.449 ± 0.036, 121 | 15.971 ± 0.061, 28 | 15.746 ± 0.075, 13 |
| (14.40–16.35) | (15.43–16.61) | (15.19–16.26) | |
| Maxillary toothrow length | 5.237 ± 0.020, 121 | 5.516 ± 0.041, 28 | 5.268 ± 0.050, 13 |
| (4.63–5.78) | (5.02–5.85) | (4.97–5.51) | |
| Interparietal width | 9.270 ± 0.044, 121 | 9.906 ± 0.079, 28 | 9.425 ± 0.102, 13 |
| (8.03–10.23) | (9.14–10.84) | (8.88–10.00) | |
| Interparietal length | 5.274 ± 0.037, 121 | 4.858 ± 0.094, 28 | 4.999 ± 0.169, 13 |
| (4.29–6.40) | (3.82–5.98) | (3.94–6.11) | |
| Parietal breadth | 14.387 ± 0.038, 121 | 13.712 ± 0.064, 28 | 13.878 ± 0.092, 13 |
| (13.26–15.50) | (12.85–14.45) | (13.21–14.60) | |
| Skull depth | 11.212 ± 0.025, 121 | 11.636 ± 0.044, 28 | 11.635 ± 0.060, 13 |
| (10.43–11.99) | (11.00–12.09) | (11.07–11.88) | |
| Least interorbital constriction/ | 27.171 ± 0.069, 121 | 24.045 ± 0.171, 28 | 24.048 ± 0.217, 13 |
| occipitonasal length ratio, × 100 (%) | (25.20–28.79) | (22.41–26.06) | (22.76–25.48) |
| Parietal breadth/ | 39.429 ± 0.100, 121 | 35.787 ± 0.250, 28 | 35.444 ± 0.300, 13 |
| occipitonasal length ratio, × 100 (%) | (36.68–41.56) | (32.63–38.94) | (33.69–37.34) |
a Mean ± standard error, sample size, and observed range (minimum–maximum) are given for external measurements,
mass, cranial measurements, and three derived ratios. Units are millimeters unless otherwise noted. La Selva (localities 18,
19), Monteverde (localities 30–32; all on the Pacific slope or crest of the Cordillera de Tilarán), and Cerro Cacao (locality
28) constituted the geographic samples (see appendix 1).
| KU |
Biodiversity Institute, University of Kansas |
| ROM |
Royal Ontario Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.