
Macrobiotus kirghizicus, Tumanov, Denis V., 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.169855 |
|
DOI |
https://doi.org/10.5281/zenodo.5629675 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87E1-D14A-FF81-8736-FADAFD17EBE1 |
|
treatment provided by |
Plazi |
|
scientific name |
Macrobiotus kirghizicus |
| status |
sp. nov. |
Macrobiotus kirghizicus View in CoL sp. nov. ( Figs. 18–23 View FIGURES 18 – 24 , 25–30 View FIGURES 25 – 33 , 34–36 View FIGURES 34 – 39 , Table 2 View TABLE 2 )
Type specimens. Holotype: slide number 195(10) (sex indet.), collected by Dr. A.V. Barabanov (Zoological Institute Russian Academy of Sciences, St. Petersburg) 17.08.1998, Kirghizia, Chui Area. Paratypes: slides 194(5, 6, 7), 198(4, 7) from the same locality. Holotype and paratypes are preserved at the Zoological Institute of the Russian Academy of Sciences, St. Petersburg, Russia. One paratype (slide number 198(7)) preserved in the collection of Binda and Pilato (Department of Animal Biology, University of Catania, Italy). Type locality: Kirghizia, Chui Area., northern slope of Kirghizian ridge, Turakaiyn canyon, vicinity of Turakaiyn village, near 2000 m a.s.l., moss from soil. Twentyeight adult and juveniles and 41 eggs (3 with developed embryos) were found.
Etymology. The name of the new species was derived from Kirghizia, its type locality.
Description. Body long and relatively plump ( Figs. 18 View FIGURES 18 – 24 , 25 View FIGURES 25 – 33 ) (body length of adult animals 478.8–685.0 m). Color white (colourless), large specimens often with granules of brown pigment. Most specimens with granular eye spots. Cuticle smooth, large specimens with well defined small areas of granulation on the ventral side. Typical distribution of these areas is the following: one area between legs I, one area between the 1st and the 2nd pairs of legs, two areas aligned in a transverse row, between legs II, three areas aligned transversally between the 2nd and the 3rd pairs of legs, one pair of areas, aligned as previously, between legs III, two rows of three areas between the 3rd and the 4th pairs of legs and a pair of areas laterally to anus. Legs with very fine dotlike sculpture (on dorsal surface and around the claw bases), better visible on legs IV.
pt of external claw length, leg IV 75.3 77.4 80.5 80.6 (75.3–88) 1.77
Ss = distance from cephalic end of buccal tube to insertion point of stylet supports.
Supplementary measurements for holotype: length of strengthening bar on buccal tube: above 33.3 m, lunules on claws of the fourth pair of legs: 8.1 x 11.8 m
Buccopharyngeal apparatus of Macrobiotus type ( Figs. 19 View FIGURES 18 – 24 , 26 View FIGURES 25 – 33 ) (for all dimensions see Table 2 View TABLE 2 ). Buccal cavity with ten scarcely visible peribuccal lamellae. Dorsal buccal armature with two granules in medial position without points and teeth ( Fig. 27 View FIGURES 25 – 33 ). Ventral buccal armature not visible. Ventral strengthening bar relatively long and exceeding half of the length of the buccal tube. Pharyngeal bulb suboval, with well developed apophyses, two macroplacoids (first with well developed constriction, longer than second) and elongated microplacoid. Between the apophyses and first macroplacoids there are thin slanting cuticular bars.
Claws extremely similar to those described by Dastych (1977) in M. adelges (junior synonym of M. ariekammensis Weglarska, 1965 ), large, with poorly developed accessory points ( Figs. 21–23 View FIGURES 18 – 24 , 28–30 View FIGURES 25 – 33 ). All claws with large lunules, especially in legs IV. Lunules of legs IV with distinct teeth, lunules of legs I–III with slightly waved edges. Legs I–III with band of thickened cuticle below claw bases.
Spherical eggs ornamented and laid free ( Figs. 34–36 View FIGURES 34 – 39 ). Egg processes consists of short domeshaped basal part and rigid spinelike apical part, clearly separated from the base with single internal septum. Without septation of apical part of processes. Basal part lacks reticulation, lateral surfaces of apical part with irregularly distributed minute spines. Bases of processes with wreath of little dark granules and two rows of light pores. The first row is located on the process base above the granules, the second row is located below the granules on the shell surface. Pores of both rows are ordered opposite each other. Shell of eggs between processes smooth, with rare, scarcely visible pores, similar to the pores at the process bases. Usually egg surface and basal parts of processes poorly visible, covered with debris particles. Diameter of eggs (without processes): 101.4–126.7 m; height of processes (about 25 on the circumference of egg): 15.2–28.7 m, their basal diameter: 8.5–13.5 m. Height of process basal part: 4.2–5.1 m.
Remarks. The new species is most similar to Macrobiotus ariekammensis . Macrobiotus kirghizicus sp. nov. differs from M. ariekammensis in the following features:
1. Two granules in the buccal armature, while M. ariekammensis has a single large triangular median tooth ( Fig. 31 View FIGURES 25 – 33 ). This character was not pointed out in the published descriptions of M. ariekammensis and M. adelges , but it is shown on a drawing by Dastych (1977, Figs. 2, 5 View FIGURES 1 – 8 ).
2. Slightly longer claws on legs IV.
3. Differences in egg morphology. Processes of M. kirghizicus eggs with clearly separated basal and apical parts, while in M. ariekammensis processes are bottle shaped or conic ( Figs. 37–39 View FIGURES 34 – 39 ). In M. ariekammensis the apical parts of the processes have internal septae and dull tips with tufts or rosettes of spines, while in M. kirghizicus spinelike apical parts of the processes lack internal septae and have only laterally distributed spines.
Macrobiotus kirghizicus sp. nov. also has some similarity in claw appearance with M. barabanovi sp. nov., but it could be easily distinguished from this species in having two macroplacoids, very different buccal armature and egg shell sculpture.
Discussion. Macrobiotus ariekammensis was included by Maucci in the tenuis group of the genus Macrobiotus . This group of species was separated on the base of the combination of the following features ( Maucci 1987a, 1987b):
1. Large dimensions of the adult animals.
2. Yellowish or light brown color.
3. Buccal tube with short strengthening bar, and with insertion point of the stylet supports placed far from the posterior end of the buccal tube.
4. Two macroplacoids, first with constriction, which could divide it in two, forming as a result three macroplacoids. Pharyngeal bulb with thin slanting cuticular bars between apophyses and first macroplacoids.
5. Long and thin claws, with well differentiated primary and secondary branches, secondary branch attached to the claw at right angle, claws IV are distinctly longer than claws I–III.
6. Large dentate lunules, especially on legs IV.
Most of the above listed characters are of small phylogenetic significance. Large dimensions, coloured body, large dentate lunules and pharyngeal bulb with two macroplacoids are widely distributed within the genus Macrobiotus . The significance of the presence of slanting cuticular bars in the pharyngeal bulb is unclear, because this structure is also present in other species of the genus Macrobiotus (for example M. macrocalix Bertolani & Rebecchi, 1993 from hufelandi group). Moreover, these structures are present in some species, belonging to the other genera of Eutardigrada (e.g. Minibiotus sp.; Tunisian population, author’s collection). Well developed baculiform structures visible on SEM photographs of buccopharyngeal apparatus of Isohypsibius saltursus Schuster Toftner & Grigarick 1977 (1978) and Dactylobiotus grandipes ( Schuster Toftner & Grigarick 1977 (1978)) ( Schuster et al. 1980) seems to be homologous to the slanting cuticular bars, visible in optical microscope. The claw length cannot be considered as a base for the grouping of species without taking into account the exact structure of the claw. Recently it was stated that details of the claw structure should be taken into consideration when discussing the taxonomic position of Macrobiotidae species ( Guidetti & Bertolani 2001). In my opinion the structure of the claws of Macrobiotus ariekammensis and M. kirghizicus is very different from the structure of the claws of other species of the tenuis group. In both species poorly differentiated main and secondary branches diverge from the very short and wide common tract. Distal part of the basal portion is very poorly marked, flattened in the frontal plane of the claw, and in the claws of the legs IV it is nearly totally fused with the stalk. On the other hand, other species of the tenuis group have different claw morphology, with well differentiated main and secondary branches, long thin common tract (secondary branch diverge at about half of the claw length) and typically developed distal part of the basal portion. Furthermore, all species of the tenuis group possesses a common feature in their claw structure – the stalk, connecting the claw base with the lunule, form a prominent frontal appendage on the claw base. This structure is absent in both Macrobiotus ariekammensis and M. kirghizicus .
Taking into account those differences in claw morphology, Macrobiotus ariekammensis and M. kirghizicus sp. nov. should not be included in the tenuis group of the genus Macrobiotus . Both species are undoubtedly closely related to each other and well detached from other species of Macrobiotus , so they should be combined into the new ariekammensis group of species.



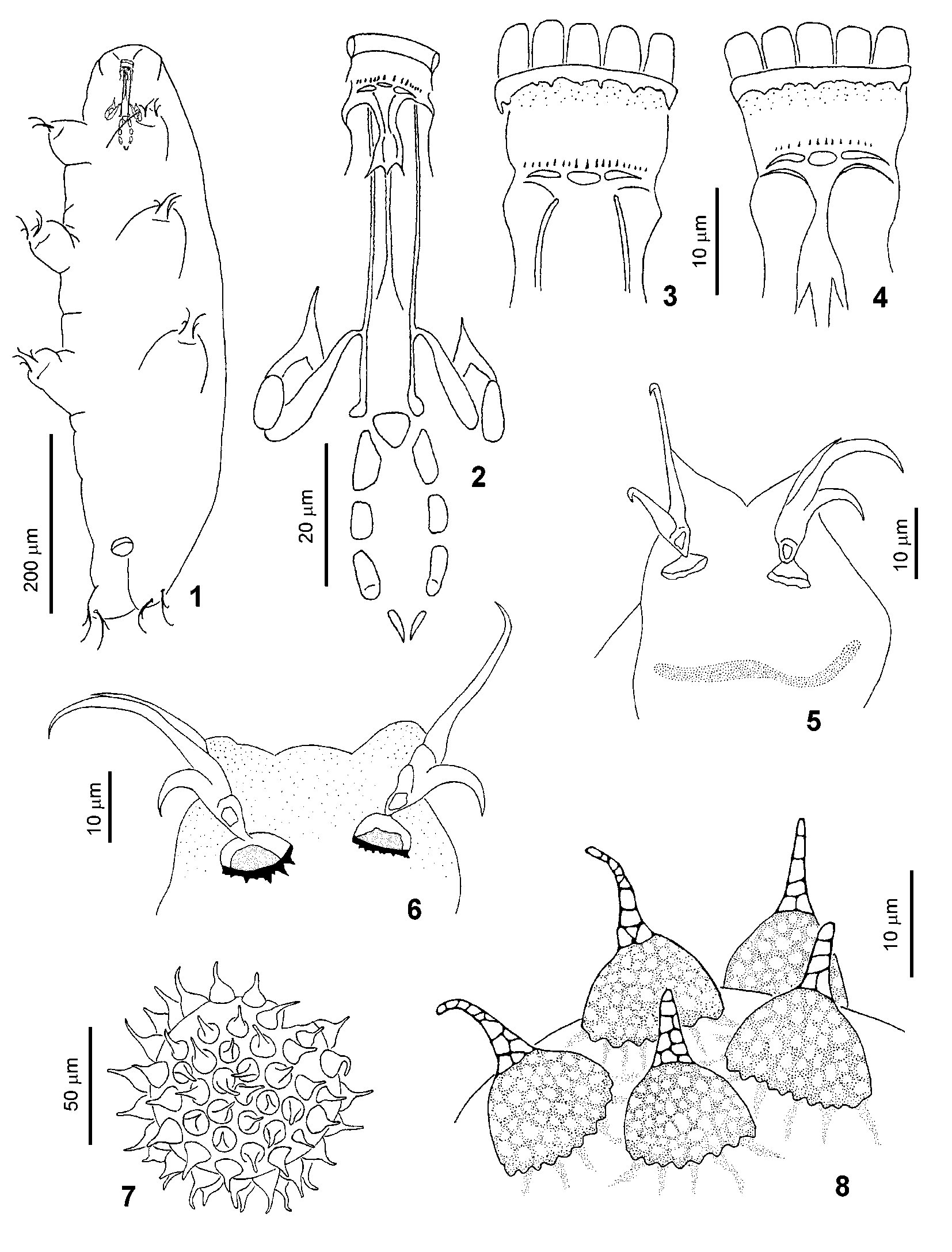
TABLE 2. Summary of morphometric data for Macrobiotus kirghizicus sp. nov. (n = 9).
| Character | Holotype | Smallest Largest Mean (Range) specimen specimen | Standard error |
|---|---|---|---|
| Body length (m) | 651.7 | 478.8 685.0 605.9 (478.8–685.0) | 22.59 |
| Buccal tube length (m) | 54.0 | 45.9 57.0 52.2 (45.9–57.0) | 1.26 |
| Ss (m) | 40.3 | 34.0 42.6 38.8 (34–42.6) | 0.94 |
| pt Ss | 74.7 | 74.2 74.7 74.3 (73.5–74.7) | 0.15 |
| Outer buccal tube width (m) | 6.7 | 6.7 7.4 6.9 (6.4–7.4) | 0.13 |
| pt of outer buccal tube width | 12.3 | 14.5 13.0 13.3 (12.3–14.5) | 0.31 |
| Placoid row length (m) | 29.8 | 25.2 33.3 28.7 (25.2–33.3) | 0.98 |
| pt of placoid row length | 55.2 | 54.8 58.4 55.3 (50.0–58.4) | 0.89 |
| Macroplacoid row length (m) | 24.4 | 20.4 26.6 24.1 (20.4–27.0) | 0.80 |
| pt of macroplacoid row length | 45.2 | 44.4 46.8 46.2 (41.7–49.7) | 0.78 |
| 1st macroplacoid length (m) | 14.1 | 11.8 14.8 13.6 (11.5–15.5) | 0.50 |
| pt of 1st macroplacoid length | 26.0 | 25.8 26.0 26.0 (21.5–28.6) | 0.63 |
| 2nd macroplacoid length (m) | 8.1 | 7.4 9.6 8.4 (7.0–9.6) | 0.32 |
| pt of 2nd macroplacoid length | 15.1 | 16.1 16.9 16.1 (14.6–17.6) | 0.36 |
| Microplacoid length (m) | 3.7 | 3.0 4.1 3.6 (3.0–4.1) | 0.16 |
| pt of microplacoid length | 6.8 | 6.5 7.1 6.9 (5.7–8.5) | 0.31 |
| External claw length, leg I (m) | 24.1 | 19.2 25.9 24.3 (19.2–26.6) | 0.76 |
| pt of external claw length, leg I | 44.5 | 41.9 45.5 46.4 (41.9–50.7) | 0.94 |
| External claw length, leg III (m) | 26.6 | 22.2 31.8 27.4 (22.2–31.8) | 1.07 |
| pt of external claw length, leg III | 49.3 | 48.4 55.8 52.8 (48.4–57.7) | 1.17 |
| External claw length, leg IV (m) | 40.7 | 35.5 45.9 42.1 (35.5–45.9) | 1.24 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |