
Oxyropsis ephippia Aquino & Sabaj Pérez, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4136.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:100BC0A4-08BF-4C1F-8E09-9DE2C5F3ADB4 |
|
DOI |
https://doi.org/10.5281/zenodo.6055888 |
|
persistent identifier |
https://treatment.plazi.org/id/03DD312C-C147-FFCD-FF42-1E92D04D75B9 |
|
treatment provided by |
Plazi |
|
scientific name |
Oxyropsis ephippia Aquino & Sabaj Pérez |
| status |
sp. nov. |
Oxyropsis ephippia Aquino & Sabaj Pérez sp. nov.
( Figs. 3–5 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Holotype: ANSP 177381, female 42.0 mm SL, Guyana, Burro Burro River ( Essequibo Drainage), Deer Falls between Deer Creek and Water Dog Falls, 4°39'53"N, 58°50'28"W, G. Watkins et al., 21 November 1997.
Paratypes (49, 22.3–44.8 mm SL).— Guyana, Essequibo –Atlantic Ocean Drainage: AMNH 265342, 3 (1 cs) 29.7–36.7 mm SL, Rupununi River 4.6 km NW of village of Massara, 3°55'34"N 59°16'49"W, M. Sabaj et al., 26 October 2002; ANSP 175918, 1, 30.8 mm SL, small creeks crossing Kurupukari–Surama River road ca. 3.0 miles from Kurupukari field station, 4°22’29”N 58°50’30”W, W.G. Saul et al., 4 February 1997; ANSP 175919, 1, 29.0 mm SL, Essequibo River approximately three hours upstream from Kurupukari field station, 4°34’17”N 58°35’17”W, W.G. Saul et al., 3 January 1997; ANSP 175920, 1, 27.0 mm SL, clearwater creek at campsite 3.1 miles from Kurupukari field station on Kurupukari–Surama River road, 4°38'0"N 58°42'59"W, W.G. Saul et al., 5 February 1997; ANSP 175921, 1, 29.5 mm SL, Essequibo River approximately two hours downstream from Kurupukari field station, 4°47'44"N 58°48'52"W, W.G. Saul et al., 20 January 1997; ANSP 175922, 4, 25.8–27.8 mm SL, Essequibo River, sandbar ca. 800 m downstream from Essequibo campsite (Maipuri), 4°45’43”N 58°45’52”W, W.G. Saul, et al., 29 January 1997; ANSP 180659, 3, 26.2–40.5 mm SL, Essequibo River (east bank) at Kurukupari, 4°39'41"N 58°40'31"W, J.W. Armbruster et al., 24 October 2002; ANSP 180662, 3, 22.3–24.7 mm SL, Rupununi River, 3.7 km SSE village of Massara, 3°51'44"N 59°17'4"W, M. Sabaj et al., 27 October 2002; ANSP 180664, 2, 29.5–35.4 mm SL, AUM 35652, 12, 22.5–30.9 mm SL, INPA 50886, 3, 27.7–30.0 mm SL, and ROM 91534, 3, 26.1–29.0 mm SL, same data as AMNH 265342; ANSP 180666, 5, 23.0–25.0 mm SL, Simoni River (trib Rupununi), four stations along river from 6.6 km SE to 3.2 km W of Karanambo Ranch, ca. 3°43'9"N 59°15'40"W, M. Sabaj et al., 29 October 2002; ANSP 180685, 1, 33.0 mm SL, Araquai Creek (trib Rupununi), 77.3 km SSE of Lethem, 2°45'45"N 59°27'60"W, M. Sabaj et al. 15 November 2003; MCP 49016, 2 alc, 26.2–30.8 mm SL, same data as ANSP 180666; MCP 49017, 2, same data as ANSP 175919.—Branco–Negro–Amazonas Drainage: ANSP 180670, 2, 40.4–44.8 mm SL, Yuora River (trib Ireng, Takutu drainage), 6.7 km NE of village of Karasabai on road to Tiger Creek village, 4°3'14"N 59°29'7"W, M. Sabaj et al., 31 October 2002.
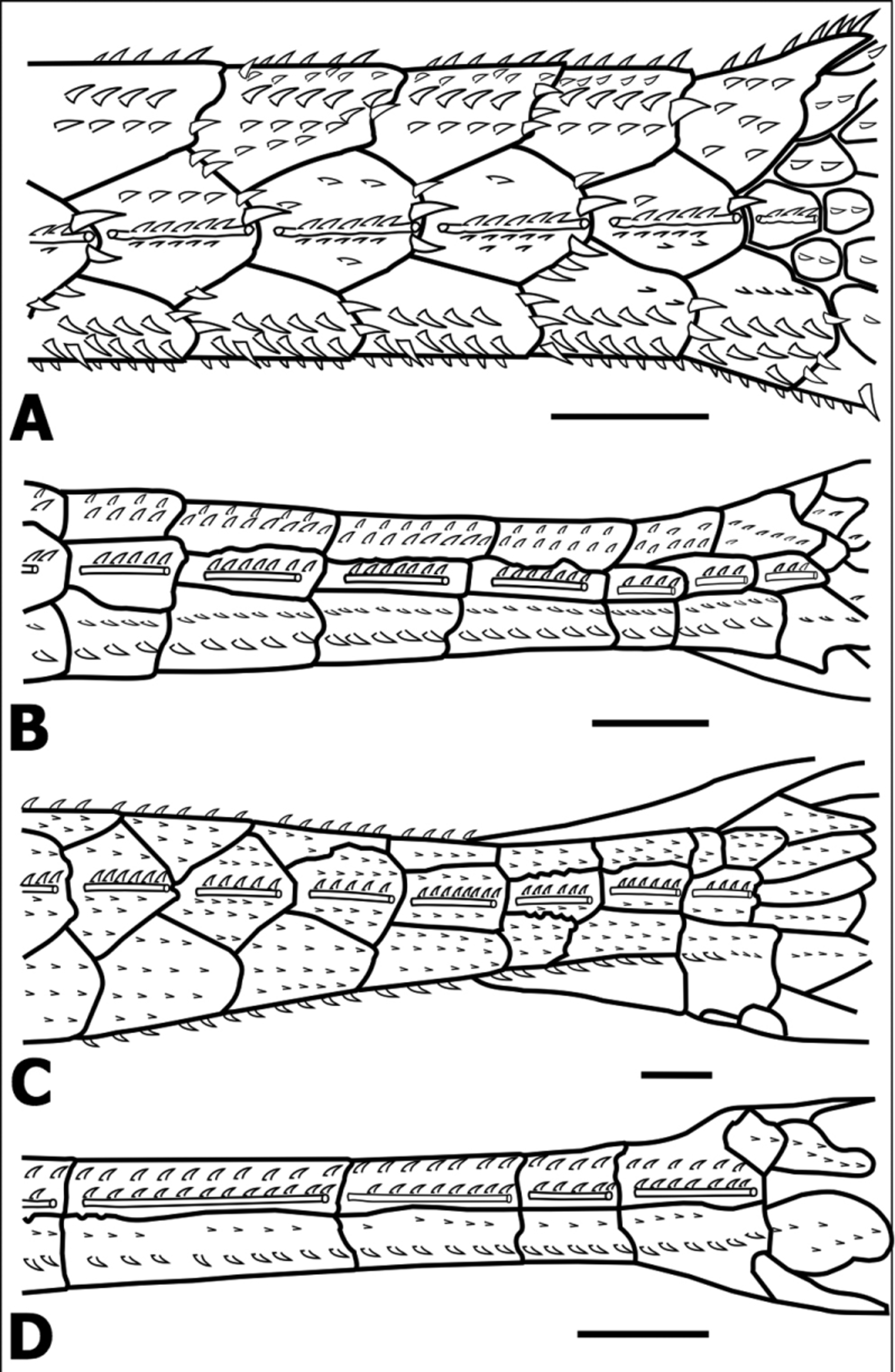
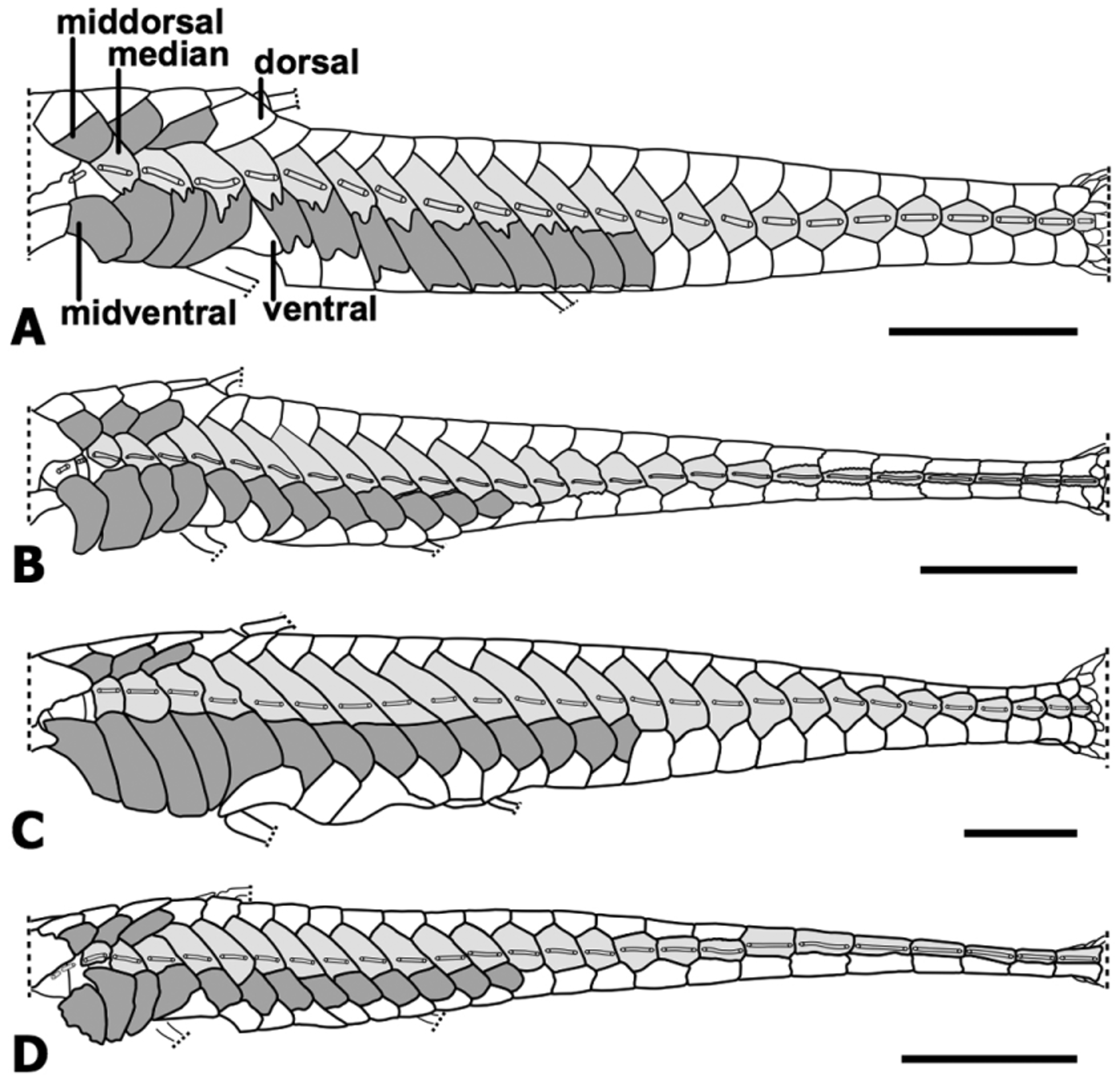
Diagnosis. Oxyropsis ephippia is distinguished from its congeners by having a keel-like row of odontodes above the lateral-line canal on each median plate that is dominated by a single odontode (one closest to posterior margin of plate) conspicuously enlarged, typically one-third to one-half larger (along base-tip axis) than preceding one in same row ( Figs. 1 View FIGURE 1 A and 4A) ( vs. keel-like row immediately above lateral-line with odontodes of roughly equal size; Figs. 1 View FIGURE 1 B–D). It is further distinguished by having the sides of the trunk below the median series, approximately between plates 7 to 12, shielded by plates of the midventral series, with the exclusion of plates from the ventral series, which are visible only in ventral view of the trunk ( Fig. 2 View FIGURE 2 A) (vs. sides of the trunk shielded by plates of the midventral series and ventral series, the latter visible in lateral and ventral views of the trunk; Figs. 2 View FIGURE 2 B–D).
Oxyropsis ephippia is further distinguished by having 23 plates along the trunk median series (vs. 18–19 in O. acutirostra and 25–26 in O. carinata and O. wrightiana ) and the caudal peduncle in cross section slightly compressed at the level of the median plates 20–23 (vs. posterior caudal peduncle depressed); from O. acutirostra by having median series of plates complete (vs. series incomplete; Fig. 2 View FIGURE 2 D); from both O. acutirostra and O. wrightiana by having a deeper caudal peduncle (mean depth 4.8% SL vs. 2.9% and 2.0%, respectively), and shorter caudal peduncle (mean length 36.0% SL vs. 42.4% and 44.4%, respectively); and from O. carinata by having a larger eye (mean orbital diameter 18.2% HL vs. 15.7% HL).
Description. General size small; largest specimen examined 48.7 mm SL. Head and trunk slender, moderately depressed (cleithral width 62.5–71.7% HL); greatest body depth at dorsal-fin origin (12.1–14.7% SL), only slightly deeper than head (11.6–12.7% SL). Dorsal profile straight to slightly convex from tip of snout to dorsal-fin origin ( Figs. 3-4 View FIGURE 3 View FIGURE 4 ). Dorsal profile of trunk straight from dorsal-fin origin to anteriormost procurrent ray of caudal fin. Ventral profile of head and abdomen approximately straight to slightly convex from snout tip to procurrent caudalfin rays. Dorsal and ventral profiles gently diverging from procurrent towards marginal rays of caudal fin. Snout smoothly rounded, parabolic in dorsal view; dorsal rostrum smooth, with paired smooth concavity in region anterior to naris. Dorsal surface between eyes and origin of dorsal fin convex in cross section, flattening caudally. Cross section of body between pectoral and pelvic-fin origins ovoid, becoming progressively fusiform-shaped (tapering posteriorly) between verticals lines through approximately tip of pectoral fin and median plates 17–18; gradually compressed at level of 6–7 most posterior median plates; CP mean width 75.5% (range 67.2–91.2%) of CP depth at plates 21–23).
Eyes moderately large, orbital diameter 17.3–19.9% HL, center of eye positioned closer to posterior tip of pterotic than to tip of snout. Eye dorsolateral on head; fourth infraorbital lateroventral. Tip of adpressed dorsal fin reaching vertical through midlength of adpressed anal-fin spine. Pelvic fin not reaching plate anterior to base of anal-fin spine. Pectoral fin reaching vertical through posterior half of pelvic-fin spine length. Serrae (teeth) along posterior aspect of pectoral-fin spine present, less developed toward basal fifth and distal fifth of spine; best developed in specimens shorter than about 27.0 mm SL, wherein serrae are conical, moderately acute, each with tip oriented toward base of spine (retrorse); serrae become less conspicuous, more blunt with increasing standard length, and tips variably slanted toward base of spine.
Dermal plates on trunk arranged in five longitudinal rows ( Fig. 2 View FIGURE 2 A): dorsal series 20; middorsal series 3; median series 23; midventral series 13 (including 4 plates between cleithrum lateral process and first plate of ventral series); ventral series typically 19-20. Ventral plates positioned either lateroventrally (1 to 4 and 11 to 20) or ventrally (5 to 10). Midventral plates deep above ventrally positioned plates in ventral series, shielding side of trunk below median series ( Fig. 2 View FIGURE 2 A). Lateral line complete, continuous along median series. Midventral plates articulating with median plates via interlocking digit-like projections; ventral and median plates in contact via overlapping smooth edges. Thoracic plates absent. Abdominal plates fully developed at 29 mm SL (development of plates incipient at about 24–25 mm SL); abdominal plates arranged in paired series of 5–9 elongate lateral plates and medial series of 4–9 polygonal plates; three or more polyhedral plates posterior to pectoral girdle; patch of small polyhedral plates between abdominal series and anal plate. Single anal plate. Canal-bearing plate between cleithrum and infraorbital bones well developed, mostly ventral except for posterior ventro-lateral portion in contact with opercle, and with lateral pore of canal close to pore of infraorbital canal between infraorbital plates 4 and 5.
Odontodes uniformly small and evenly distributed dorsally on head, progressively arranged in longitudinal rows toward sides and dorsum of trunk. Subtle keel-like longitudinal row of odontodes immediately above lateralline canal on each median plate is dominated by single odontode (close to posterior plate margin above pore of lateral-line canal) that is conspicuously enlarged, typically one-third to one-half longer (along base-tip axis) than preceding one in same row ( Fig. 1 View FIGURE 1 A); keel-like row most noticeable in dorsal view of trunk between median plates 8 and 18. Enlarged odontode usually paired with similarly enlarged odontode below lateral-line canal.
Midventral plates 8 to 13 with area of slightly enlarged odontodes, particularly along margin of plates; odontodes less noticeable with increasing SL. On ventral side of body, odontodes tightly and evenly arranged over ventral canal-bearing plates, abdominal plates, anal plate, and ventral series of plates posterior to anal plate. Odontodes on ventral margin of pelvic-fin spine enlarged; odontodes on ventral margin of pectoral-fin spine moderate in size.
Anterior tip of rostral plate with narrow pad of soft tissue; odontodes dorsal and ventral to tip irregularly arranged. Anterior and lateral rostral snout plates not noticeably reflected ventrally. Nuchal plate width about 2.5 times width of dorsal-fin spine at origin.
Teeth small, slender; premaxillary teeth 19–27, mandibular teeth 16–26. Oral disk round, papillose. Maxillary barbels present, short. Dorsal fin i,7; pectoral fin i,6; pelvic fin i,5, anal fin i,5. Adipose fin absent.
Sexual dimorphism. Male genital papilla present. In males, odontodes small and tightly packed on plates 2–4 of trunk ventral series, lateral to anal plate and anus region; odontode arrangement more spaced on anal plate and on plates between anal fin and anus. In females, distribution of odontodes on same ventral plates not noticeably different from patterns on adjacent plates.
Coloration. Ground color in ethanol tan ( Figs. 3–4 View FIGURE 3 View FIGURE 4 ). Dark brown punctiform epidermal melanophores in small clusters irregularly distributed on dorsal and lateral portions of head ( Fig. 4 View FIGURE 4 ), sometimes leaving pale middorsal region on snout ( Fig. 3 View FIGURE 3 ). Dark brown deep-lying punctiform melanophores roughly arranged in six middorsal blotches (typically, one anterior to dorsal fin, one at base of dorsal fin, and four posterior to dorsal fin). Brown midlateral stripe through cleithrum lateral process, along trunk median and midventral series of plates to base of caudal fin; stripe more diffuse posteriorly, variably connected to darkened lanceolate plates at base of caudal fin. Dorsal portion of cleithrum lateral process, in contact with first midventral plate, dark brown.
Ventral surface of head and trunk, between posterior lip and anus, pale, scarcely pigmented. Punctiform melanophores concentrated on anterior lip, extended laterally to barbels; scattered at bases of anal, pectoral and pelvic fins. Uneven sprinkling of deep-lying dark melanophores over ventral canal-bearing plates and along ventral series of plates.
Caudal-fin with 6–7 vertical bands of brown melanophores restricted to fin rays; proximal bands variably coalesced into darker, slightly asymmetrical blotch on basal lower half of caudal fin; basal blotch variably connected with dark lanceolate plates ( Fig. 5 View FIGURE 5 A). Other fins with faint dark bands along anterior edge of spine, continuous with diffuse bands on branched rays (roughly, 8–9 bands on dorsal fin, 3–4 on anal fin, five on pectoral fin, and three on pelvic fin). Interradial membranes of all fins clear, unpigmented.
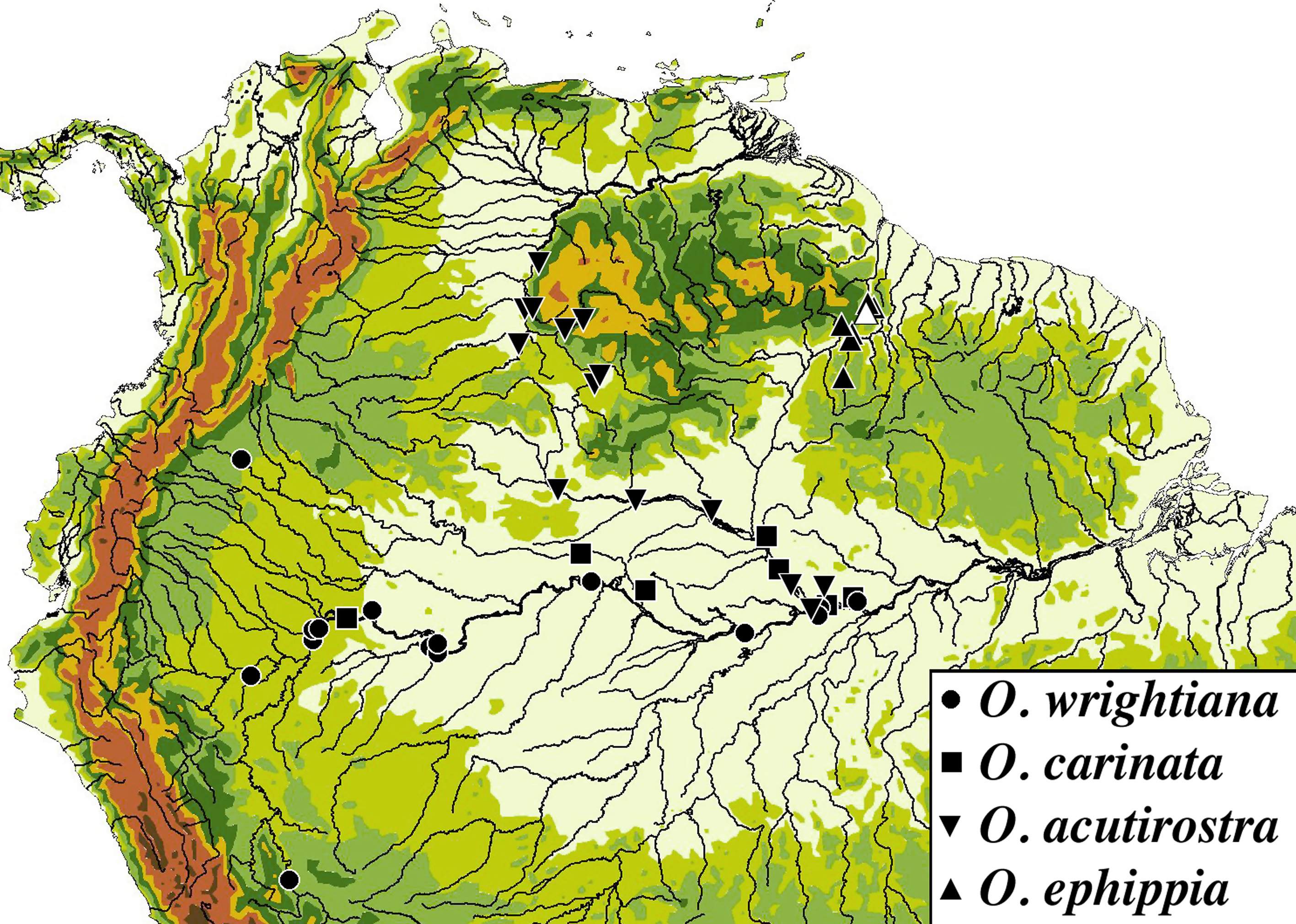
Distribution. Oxyropsis ephippia is recorded from Guyana ( Fig. 6 View FIGURE 6 ) on both sides of the shallow divide separating the Essequibo watershed (Atlantic Drainage) from the Branco (Negro–Amazonas Drainage). In the Essequibo Basin, it is known from the mainstem Essequibo, and two of its left bank tributaries (Burro Burro, Rupununi, and their tributaries). In the Branco Basin it is know from the Yuora River, a small tributary of the Ireng draining the southeastern limits of the Pakaraima Mountains.
The cichlid Guianacara dacrya described by Arbour & López–Fernández (2011) occurs in many of the same streams (e.g., Burro Burro, Rupununi and Yuora), and shares a similar distribution pattern on both sides of the Essequibo –Branco divide. The two watersheds are intermittently connected in wet years (between May and September) when the Rupununi savannas between the Pakaraima and Kanuku mountains becomes a vast flooded plain known as Lake Amaku (Lowe– McConnell 1964).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |