
Archiinocellia Handlirsch, 1910
|
publication ID |
https://doi.org/10.11646/zootaxa.4951.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:557825A0-714A-426A-917F-1C9AB7372C30 |
|
DOI |
https://doi.org/10.5281/zenodo.4681527 |
|
persistent identifier |
https://treatment.plazi.org/id/03DE878E-FFF9-C45E-FF67-FD29C9FB8435 |
|
treatment provided by |
Plazi |
|
scientific name |
Archiinocellia Handlirsch, 1910 |
| status |
|
Genus Archiinocellia Handlirsch, 1910 View in CoL , sit. nov.
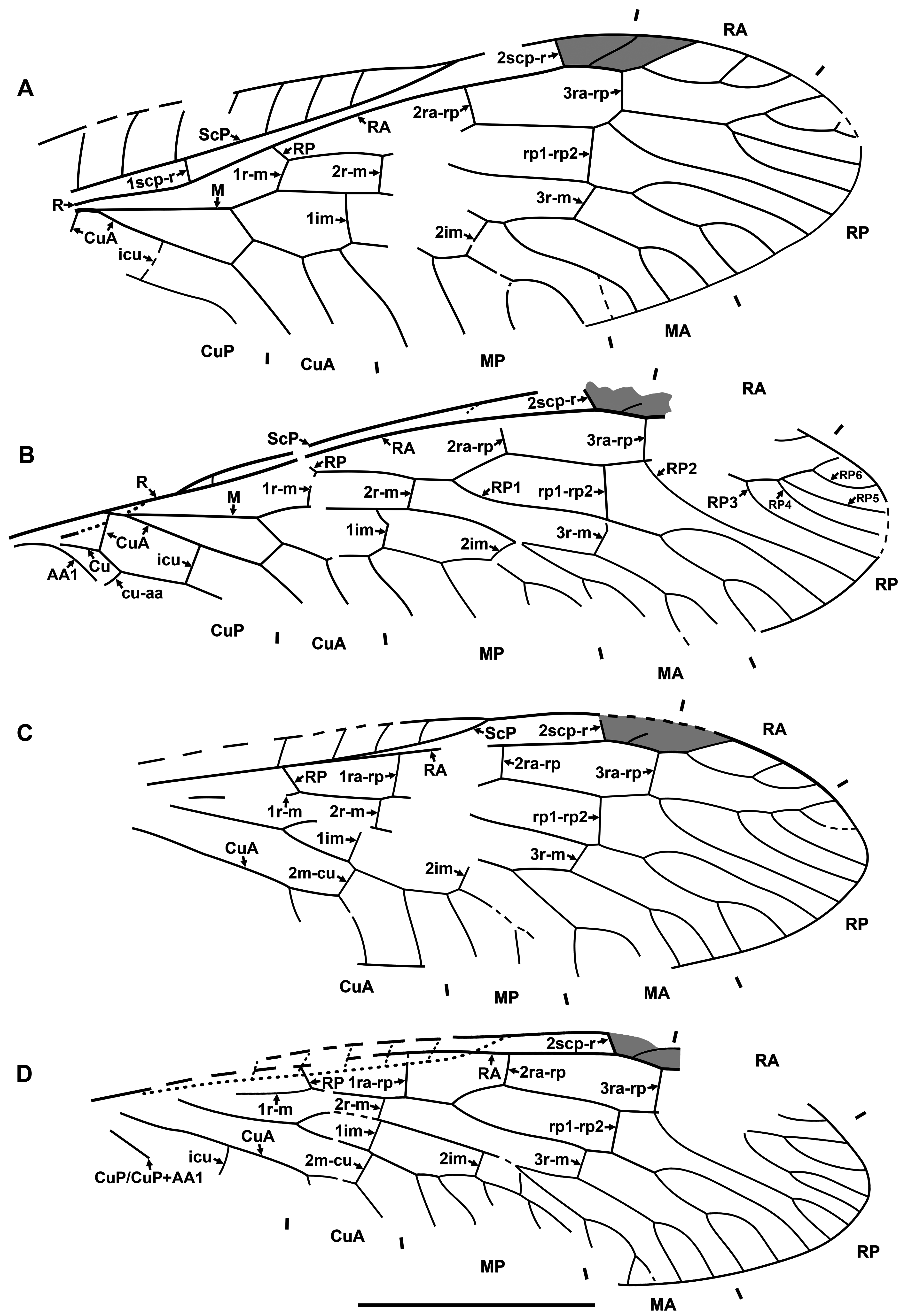
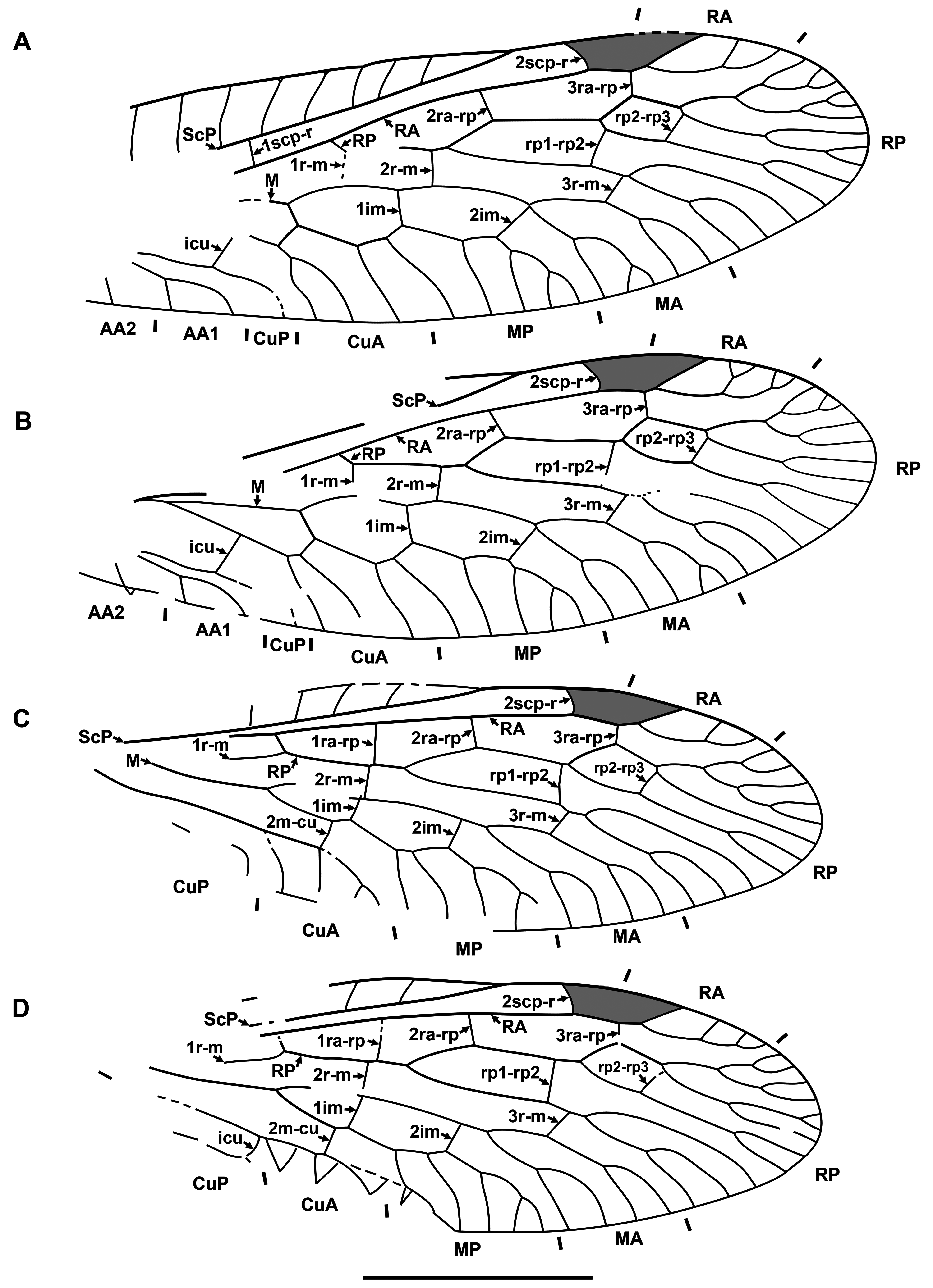
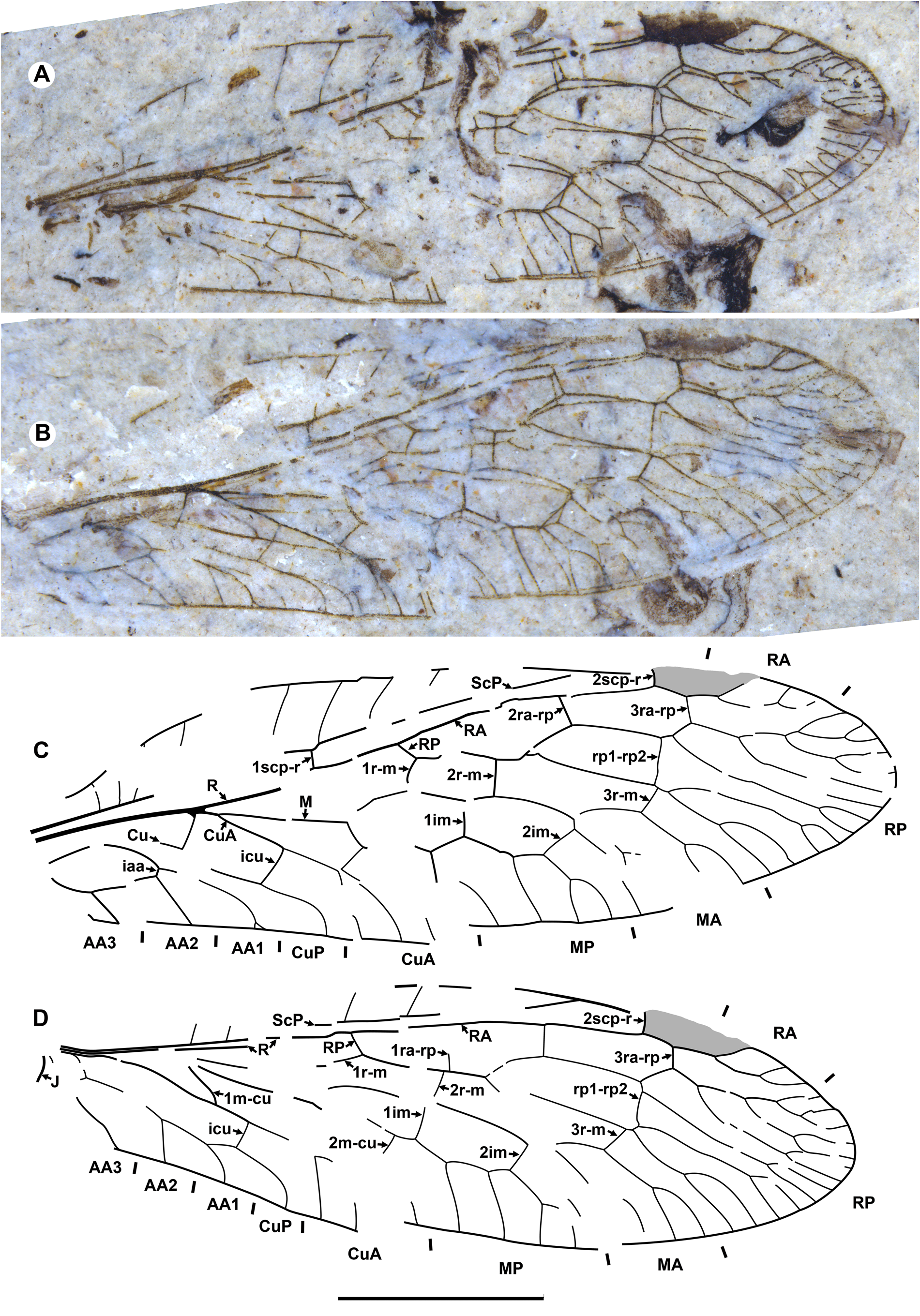
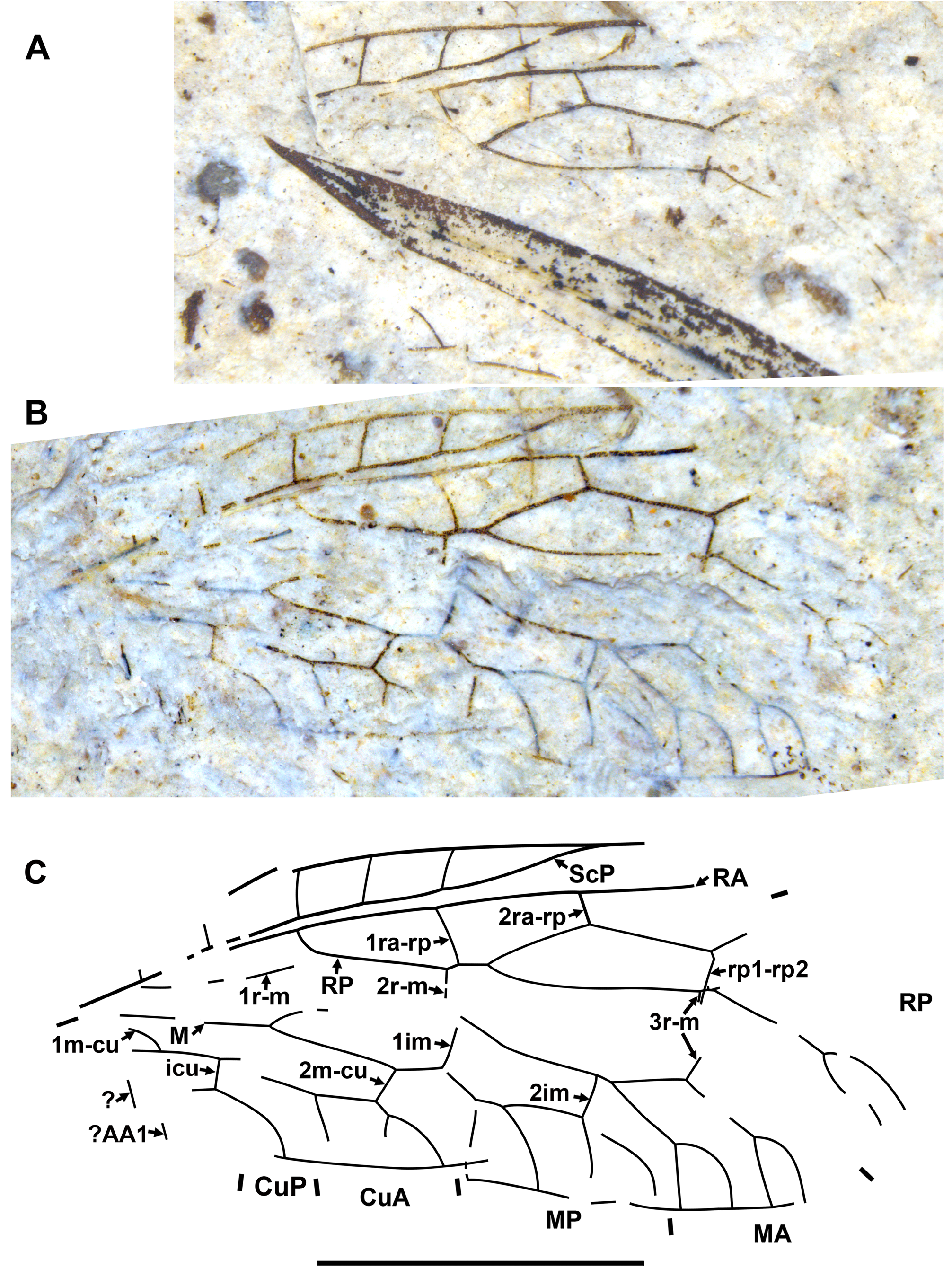
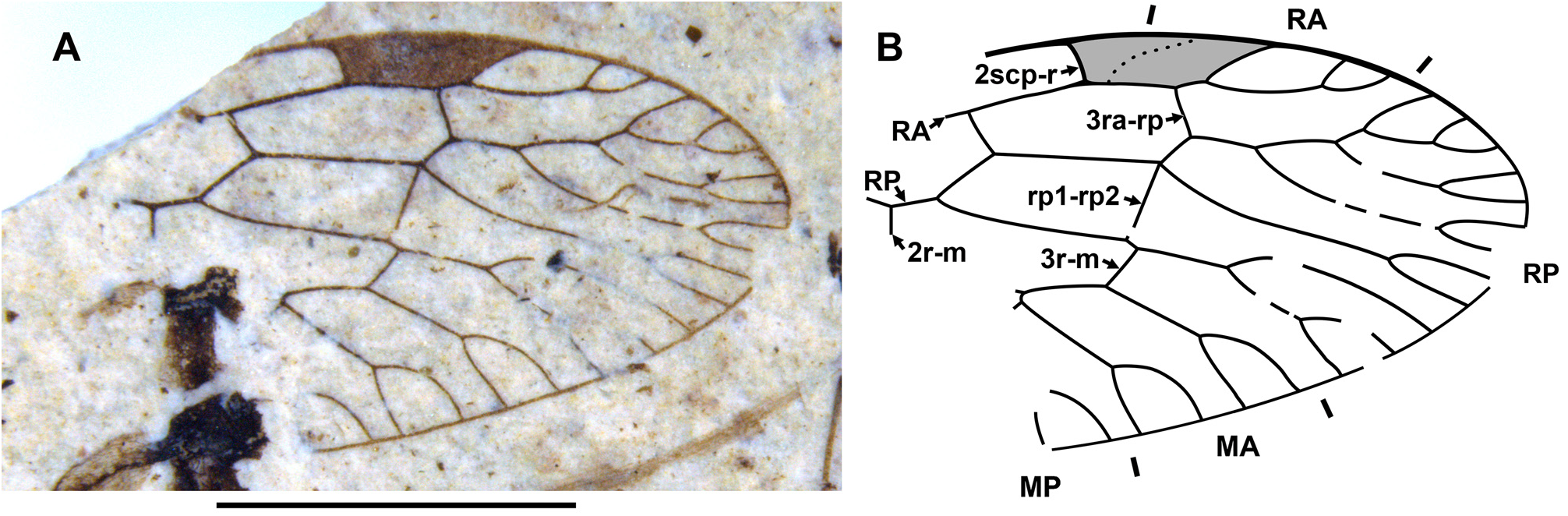
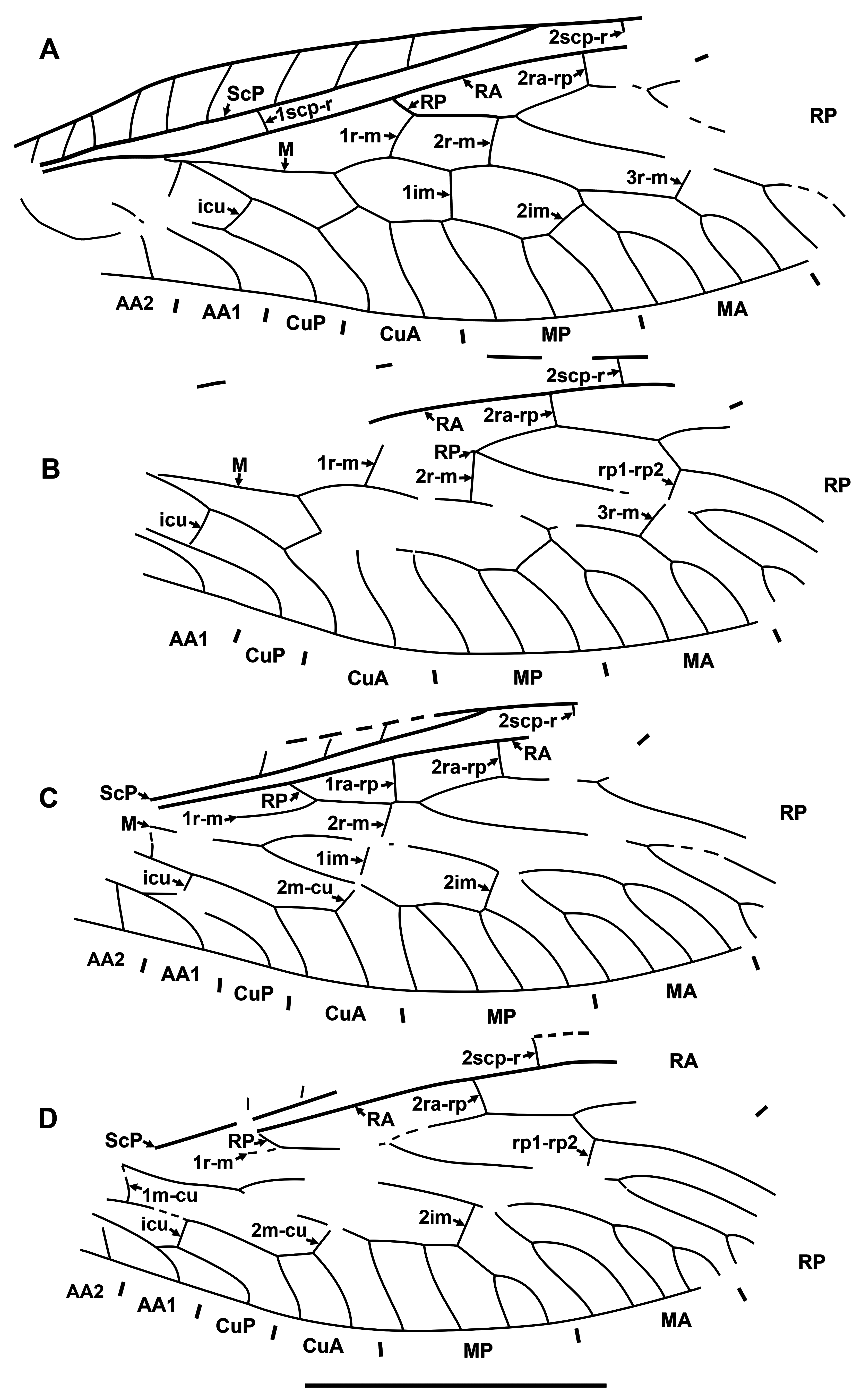
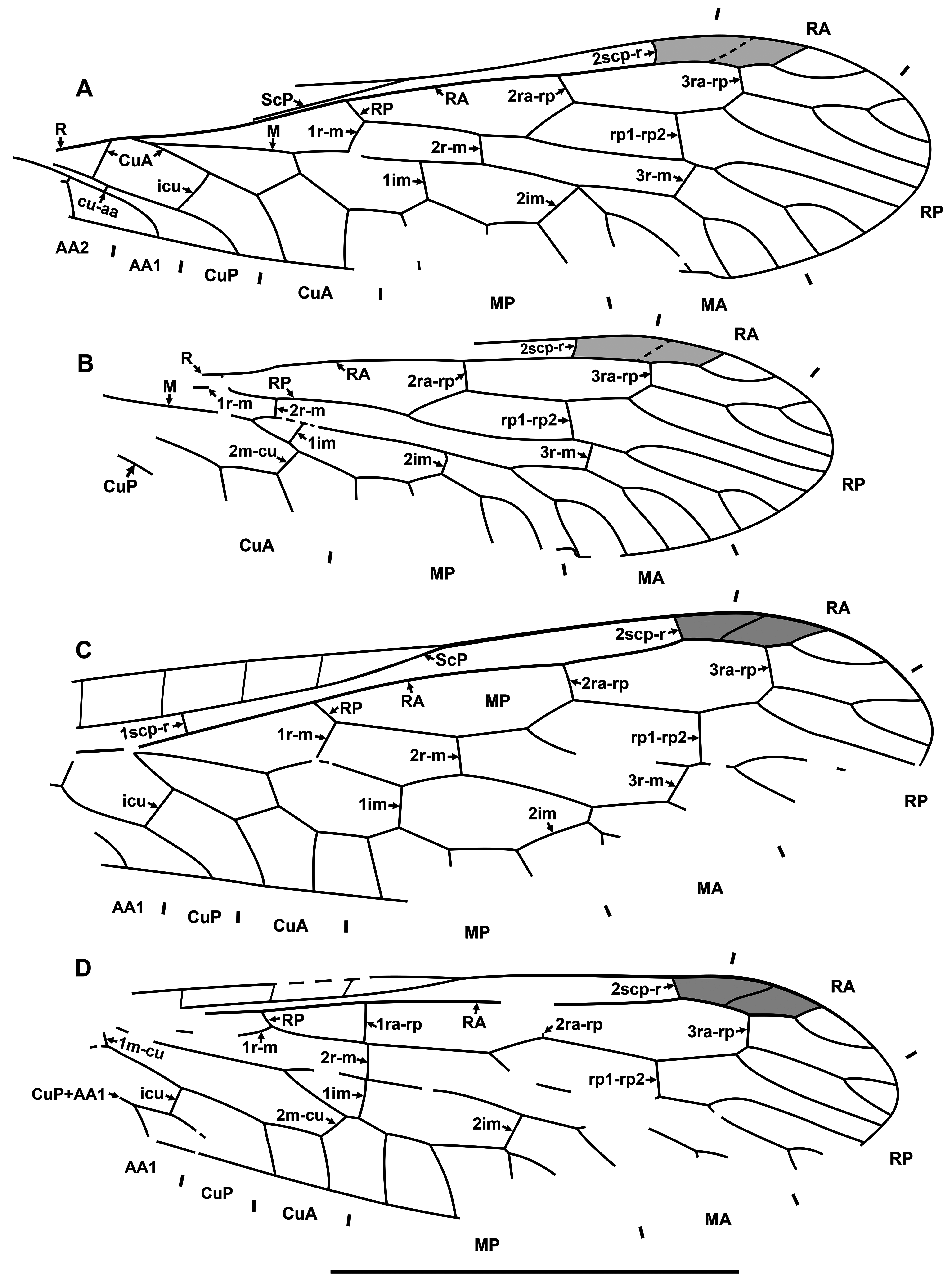
Diagnosis. Relatively small raphidiids (forewing up to 8 mm long), distinguished from other genera of the family by the following combination of character states. Fore- and hind wings: (1) ScP short (length from termination to pterostigma greater than that of pterostigma) [relatively long ScP (length from termination to pterostigma approximately equal to or less than that of short pterostigma) in Megaraphidia ]; (2) pterostigma elongate (4.5–5.1 times longer than width measured along to costal margin) [short in Megaraphidia , Archiraphidia ]; (3) 3ra-rp located within the pterostigma; (4) RP1 twice forked, proximal fork proximad level of 3ra-rp. Hind wing: (5) two doi [one in Archiraphidia ]; (6) CuA with one branch [three in Florissontoraphidia].
Type species. Archiinocellia oligoneura View in CoL , by monotypy.
Species included. A. oligoneura View in CoL , A. protomaculata ( Engel, 2011) , comb. nov.
Occurrence. Ypresian of the Okanagan Highlands at Horsefly, British Columbia, Canada, Canada; the late Ypresian Green River Formation, Colorado, United States of America.
Remarks. The genus was originally considered to be of unknown family ( Handlirsch, 1910; Martynov, 1925; Carpenter, 1936). Aspöck et al. (1991) assigned it to the Inocelliidae , but more recent authors have treated it again as Raphidioptera incertae sedis ( e.g., Oswald, 1990; Carpenter, 1992; Nel, 1993; Engel, 2002, 2009). Here, we assign it to the Raphidiidae by venation characteristic of the family, including the placement of crossvein 2ra-scp at the proximal end of the pterostigma.
Archiinocellia protomaculata from the Green River Formation was originally assigned to Agulla ( Engel, 2011: Figs 1–9 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ). We believe that this is incorrect by its wing venation, which possesses at least three important character states not present in that genus. First, the basal 1r-m in the hind wing of this species is long and subparallel to R (clearly visible in Fig. 17D View FIGURE 17 and the paratype USNM 31752), but it is crossvein-like in all known species of Agulla (see e.g., Carpenter, 1936: Fig. 1 View FIGURE 1 ). Secondly, the crossvein 3ra-rp in both wings is located within the pterostigma in this species, whereas it is normally located distad the pterostigma in all known species of Agulla ( e.g., see Carpenter, 1936: Fig. 1 View FIGURE 1 ); very rarely, adventitiously at the distal end of pterostigma ( e.g., Wognum, 1959: Figs 20, 35; Aspöck, 1987: Fig. 1 View FIGURE 1 ); and only abnormally within the pterostigma in one of the wings ( e.g., Wognum, 1959: Fig. 21). Thirdly, ScP is very short in this species, whereas it is long in all species of Agulla (from its termination to the pterostigma is nearly equal to the length of pterostigma or slightly shorter in the species of Agulla ).
We assigned Archiinocellia protomaculata to this genus as its venation is very similar to the preserved venation of A. oligoneura , agreeing with all character states of the genus diagnosis.
Of extant genera, the venation of Archiinocellia protomaculata is most similar to that of Alena Navás, 1916 , the southern-most genus of Raphidiidae in the Western Hemisphere, occurring in south-western United States of America and Mexico (south to Chiapas) with ten species ( Aspöck, 1975; Aspöck & Aspöck, 2013). A combination of the following features is characteristic of only five species of this genus among extant Raphidiidae : (A) the basal crossvein 1r-m in the hind wing is long, subparallel to R; (B) very short ScP (from its termination to pterostigma is much longer than the length of pterostigma), and (C) the proximal position of the crossvein 3ra-rp ( i.e., located within the pterostigma) in both wings. The venation of A. protomaculata shares all of these character states. It is most similar to that of some species of the Mexican genus Alena ( e.g., Alena americana Carpenter, 1958 , A. caudata ( Navás, 1914b) ; see Carpenter, 1958: Fig. 1 View FIGURE 1 ; Aspöck & Aspöck, 1970: Fig. 3 View FIGURE 3 ). The only venational feature clearly distinguishing A. protomaculata from extant species of Alena is the branching of RP1. It is forked twice, with the proximal fork located very deeply, proximad the level of 3ra-rp, while RP1 is forked once in most species of Alena , and in the few other species of Alena it is forked twice ( i.e., A. americana , A. tenochtitlana Aspöck & Aspöck, 1978 ), the proximal fork is relatively shallow (distad the level of 3ra-rp). However, the shape of the male terminal segments of Archiinocellia protomaculata fundamentally differs from those of all species of Alena . In A. protomaculata , the ninth tergite extends laterally to the ventral margin, and is probably fused with the ninth sternite to form a ring, a plesiomorphic condition at the family level, characteristic of all Raphidiidae except Alena , in which these are widely separated (Haring et al., 2010). The apparent absence of sclerotized gonocoxites 9 in A. protomaculata is also noteworthy and probably plesiomorphic at the family level. But we do not know if these terminal characters are also shared by other two species of the genus.
In all extant genera, these are the largest sclerites of the terminalia and are strongly sclerotized ( Aspöck & Aspöck, 2008). Gonocoxites 9 are also well developed in the Baltic amber Succinoraphidia exhibens Aspöck & Aspöck ( Aspöck & Aspöck, 2004: Fig. 6 View FIGURE 6 ).
Makarkin & Archibald (2014) referred Archiraphidia ? somnolenta ( Scudder, 1890) from Florissant to that genus only provisionally. It is similar to Archiinocellia in some ways, sharing with Archiinocellia oligoneura the following character states: (1) Archiraphidia ? somnolenta has a long pterostigma, 4.5 to 5 times longer than wide ( A. oligoneura , 4.6–4.7), while those of Archiraphidia species are much shorter, 2.3 to 3.2 times longer than wide, and (2) both have two doi in the hind wing (one in species of Archiraphidia ). However, the ScP of Archiraphidia ? somnolenta is slightly longer than in Archiinocellia oligoneura and A. protomaculata , somewhat differing from character state (1) of the Archiinocellia diagnosis, and the proximal-most branching of RP1 is clearly distad the level of 3ra-rp, contradicting diagnostic character state (4). The taxonomic placement of Archiraphidia ? somnolenta will be clarified by the discovery of more complete specimens; the species does not belong to Archiinocellia and probably does not belong to Archiraphidia .
Of fossil genera, the venation of Archiinocellia is most similar to that of Archiiraphidia and Florissontoraphidia, which share character states A–C. However, Archiinocellia clearly differs from these genera as stated in its diagnosis.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Raphidiomorpha |
|
Family |