
Caenotropus schizodon, Scharcansky, Alexandre & Lucena, Carlos Alberto Santos De, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.178283 |
|
DOI |
https://doi.org/10.5281/zenodo.5695383 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF87F1-FFE4-FF98-FF09-FC34FCBEF836 |
|
treatment provided by |
Plazi |
|
scientific name |
Caenotropus schizodon |
| status |
sp. nov. |
Caenotropus schizodon View in CoL , new species
( Fig. 1 View FIGURE 1 ; Table 1 View TABLE 1 )
Caenotropus labyrinthicus, Vari et al. 1995: 26 View in CoL (in part, MZUSP 29357).
Holotype: Brazil, Mato Grosso: Rio Arinos, Porto dos Gaúchos, 11º31´51”S, 57º25´08”W, R. Reis et al., 19 Jan 2002, MCP 29971 (71.53 mm SL).
Paratypes: All from Brazil. Mato Grosso: Rio Peixoto de Azevedo along BR-163, near Peixoto de Azevedo, 10º12`54”S, 54º58`04”W, R. Reis et al., 23 Jan 2002, MCP 30101 (4, 75.86–82.03 mm SL, one specimen, 75.86 mm SL, c&s; one specimen, 81.84 mm SL, SEM); Rio Teles Pires on road MT-220 between Sinop and Porto dos Gauchos, 11º39´15”S, 55º42´8”W, R. Reis et al., 20 Jan 2002, MCP 30103 (22, 23.42–90.52 mm SL, two specimens, 25.41 and 28.11 mm SL, SEM). Pará: Rio Tapajós, Pederneiras, Michael Goulding, 24 Oct 1983, MCP 16953 (1, 48.86 mm SL, c&s); same locality, MZUSP 29357 (13, 55.23– 46.12 mm SL).
Diagnosis. Caenotropus schizodon is readily distinguished from C. labyrinthicus and C. mestomorgmatos by its bifid premaxillary ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ) teeth vs. unicuspid teeth that are pointed or blunt distally. Additionally, C. schizodon differs from C. labyrinthicus by its distinct midlateral stripe extending from the snout to the base of the caudal fin vs. a diffuse midlateral stripe in most individuals of the latter species. Caenotropus schizodon differs from C. maculosus by the absence of teeth in lower jaw vs. the presence of dentition in most individuals, in having the distal portions of the anterior distal-fin rays dusky vs. having a distinct patch of dark pigmentation in that region, and 28–30 (n=18, m=29.0) lateral-line scales vs. typically 27, rarely 28.
Description. Body relatively robust, more so in larger individuals. Greatest body depth at dorsal-fin origin. Dorsal profile of head distinctly convex anteriorly, straight to slightly convex from region proximate to vertical through posterior nostril to rear of supraoccipital. Dorsal profile of body straight to slightly convex from rear of head to dorsal-fin origin, straight and posteroventrally slanted along dorsal-fin base, and slightly convex to slightly concave from base of last dorsal-fin ray to caudal peduncle. Dorsal surface of body with transversely obtuse median ridge extending five to six scales anterior of dorsal fin. Dorsal surface of body smoothly rounded transversely posterior of fin. Ventral profile of body more or less gently convex from tip of lower jaw to caudal peduncle. Prepelvic region transversely flattened with obtuse lateral angles and median series of scales. Postpelvic region gently rounded posterior to anus.
Dorsal-fin rays iii, 9 (n=18). Fin margin ranging from slightly convex to straight.
Anal-fin rays ii, 7–8 (7, n=15; 8, n=3). Fin emarginate, three anterior branched rays more elongate than last three or four rays. Pectoral-fin rays i, 11–14 (11, n=4; 12, n=12; 13, n=1; 14, n=1). Fin profile asymmetrically pointed with third or fourth ray from dorsal margin longest. Fin extends to or nearly to pelvic-fin insertion. Pelvic-fin rays i, 8. Fin distal margin somewhat convex, with first branched ray longest; fin reaches posteriorly about two-thirds distance to anal fin. Caudal fin forked, upper lobe slightly longer in some individuals; most individuals with damaged tips. Adipose fin present. Head profile distinctly rounded anteriorly, but pointed overall; upper jaw fleshy and somewhat longer than lower jaw; anterior portion of lower jaw moderately fleshy. Mouth barely inferior; maxilla extending posteriorly slightly beyond vertical through posterior nostril. Nostrils proximate. Adipose eyelid present, more developed in larger individuals, with vertically ovoid opening center of eye. Posterior margin of subopercle pointed. Series of small bifid brown-tipped teeth present in upper jaw. No lower jaw teeth present in any examined specimen. All lateral-line scales pored, canals straight. Arrangement and relative sizes of anterior lateral-line scales somewhat irregular, with sixth scale distinctly smaller. Last lateral-line scale tubular.
Color in alcohol. Ground coloration tan. Anterior and dorsal portions of fleshy upper lip and dorsal surface of head darkly pigmented. Body with distinctive midlateral stripe extending from just posterior to orbit to caudal peduncle and cross middle caudal-fin rays in all specimens of all examined sizes. Dorsal surface of body above midlateral stripe brown. Region ventral of stripe yellow or light brown. Rotund to horizontallyelongate spot of dark pigmentation overlying midlateral stripe always present about five scales posterior of opercle. One, rarely two, horizontal series of somewhat very diffuse dark patches below midlateral stripe body. Each dark patch situated in region of overlap successive scales in horizontal series. Three, sometimes four, patches of dark pigmentation above midlateral stripe more obvious at all sizes than those ventral to midlateral stripe. Margins of anterior dorsal-fin rays outlined by small chromatophores, with broad patch of dark pigmentation on membranes of unbranched and first to fifth branched rays. Central and basal portions of membranes of unbranched and first to second rays hyaline. Basal portions of posterior fin membranes dusky. Adipose fin light colored with few chromatophores. Anal and paired fins hyaline. Caudal-fin rays dusky.
Distribution. Upper and middle portions of Rio Tapajós basin, Rio Amazonas basin.
Etymology. The specific name, schizodon , from Greek words schize for divided and odons teeth, is in reference to the bifid premaxillary teeth present in the species.
Comments. Vari et al. (1995), following Géry (1964), synonymized the genus Tylobronchus Eigenmann (1912) (type-species: T. maculosus ) with Caenotropus . The main argument was the wide variation they found in different populations with respect to the presence of teeth in the lower jaw, a principal diagnostic character given by Eigenmann to differentiate his new genus from Caenotropus . Although the proposed synonimization is not invalided, the authors overlooked Eigenmann’s statement (1912: 271; 272) about the presence of bifid premaxillary teeth in Tylobronchus (= Caenotropus ) maculosus , which is an important differential character for the species. Recognition of Tylybronchus would render Caenotropus non-monophyletic.
Relationships. Vari (1983) pointed out a series of characters diagnostic for Chilodontidae along with apomorphic characters for Chilodus and Caenotropus . Subsequently, Vari et al. (1995: 10–12) expanded the study of interrelationships in the family and identified eight synapomorphies (45 to 52) for Caenotropus . Cleared and stained specimens of C. schizodon were examined in order to determine whether they share the synapomorphies for Caenotropus and the position of the species in the interrelationships proposed by Vari et al. (1995). The reader is referred to Vari et al. (1995) for further details on these characters and conditions in outgroup taxa.
The characters, all translated from Vari et al. (1995) - the well-developed process on the lateral surfaces of pterotic anterodorsal to the pterotic articular process (46), the very well-developed canal system in the middle and dorsal portions of the angulo-articular (47), the dorsal extension of the supracleithrum along the posterior margin of the posttemporal (49), the curved, anteriorly shifted postcleithrum (51), and the horizontal elongation of the terminal scale of the lateral-line series (52) - are common to C. schizodon and its congeners. The others three characters, 45, 48, and 50 require additional discussion.
Character 45 of Vari et al. (1995) is the reduction of the sixth infraorbital to an ossified tube that surrounds the associated laterosensory canal. Caenotropus schizodon has a pronounced laminar bone ventrally associated with the laterosensory canal segment. Such a laminar bone occurs in proximate outgroups of the Chilodontidae and Chilodus (see Vari et al., 1995: fig. 1).
Character 48 of Vari et al.(1995), is the pronounced development of the anterior process of the metapterygoid resulting in the elimination of the metapterygoid-quadrate fenestra, and the development of an interdigitating joint between the anterior metapterygoid process and the proximate portion of the quadrate. Caenotropus schizodon shows different degrees of this character. While the character is present in MCP 16953 (48.86 mm SL), in another specimen examined (MCP 30101; 75.86 mm SL) the metaperygoid extends to near the quadrate, but there is no contact between them and, in consequence, no interdigitating joint exists. It may be considered an ontogenetic difference.
Character 50 proposed by Vari et al. (1995) is the posteriorly directed process that arises from the posteromedial surface of the dorsal portion of the supracleitrhum. Caenotropus schizodon lacks this character. The results above indicate that characters 45 and 50 are homoplastic conditions and parsimoniously interpreted as autapomorphies not exclusive to C. schizodon .
Va ri et al. (1995) have proposed that C. maculosus is most closely related to C. labyrinthicus . In this analysis, including C. schizodon and the bifid dentition character, C. schizodon would be the sister species of C. labyrinthicus . The presence of teeth in the lower jaw (59), the spot of dark pigmentation within the midlateral stripe on the body (60), and the reduction of the dark pigmentation in the dorsal fin (61), traits that were considered autapomorphics of C. labyrinthicus by Vari et al. (1995), are now reinterpreted as synapomorphies defining the lineage formed by those two species. The hypothetized clade C. maculosus , C. schizodon , and C. labyrinthicus is defined by 3 synapomorphies: bifid teeth (regression to a primitive condition in C. labyrinthicus , hence simple teeth is autapomorphic to this species) and characters 54 and 56 of Vari et al. (1995), respectively, the posterior extension of the subopercle into a distinct process extending posterior of the limit of the opercle and the ossified laterosensory tube between the middle caudal-fin rays posterior of the elongate terminal scale of the lateral line ( Fig. 4 View FIGURE 4 ).
Posterior termination of the dorsal-fin base to origin 18 28.0 26.6 30.6 28.3 0.97 of adipose fin
We are grateful to Osvaldo Oyakawa (MZUSP) and Helmut Wellendorf (NMW) for the loan and information about specimens in their care, Richard Vari, U.S. National Museum of Natural History, for information on C. mestomorgmatos and corrections and suggestions on the manuscript, and to the staff of the Centro de Estudos em Microscopia Eletrônica e Microanálises (CEMM PUCRS) for assistance. The photograph of the holotype, Fig. 1 View FIGURE 1 , was taken by Aloísio Braun. Financial aid for the expedition where the new species was collected was provided by the PRONEX project “Conhecimento, conservação e utilização racional da diversidade da fauna de peixes do Brasil ” FINEP/CNPq grant 661058/1997-2).


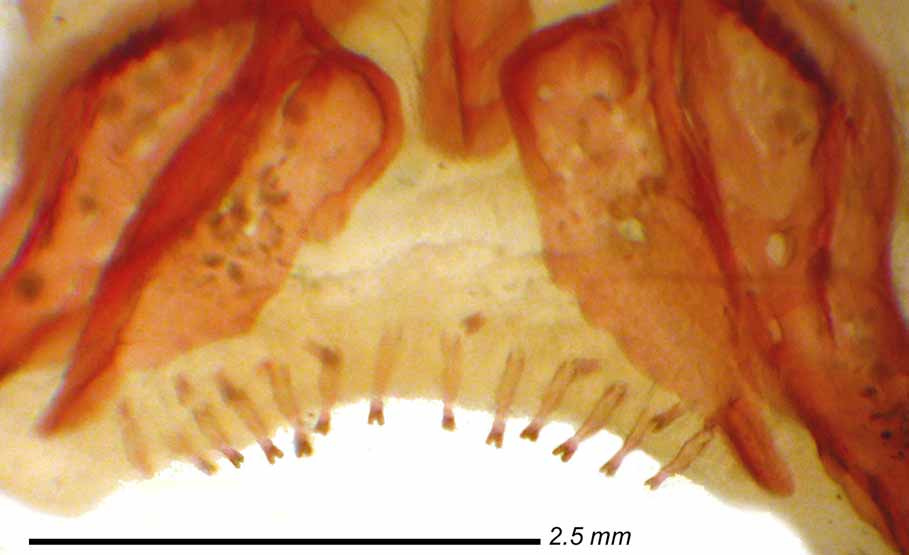
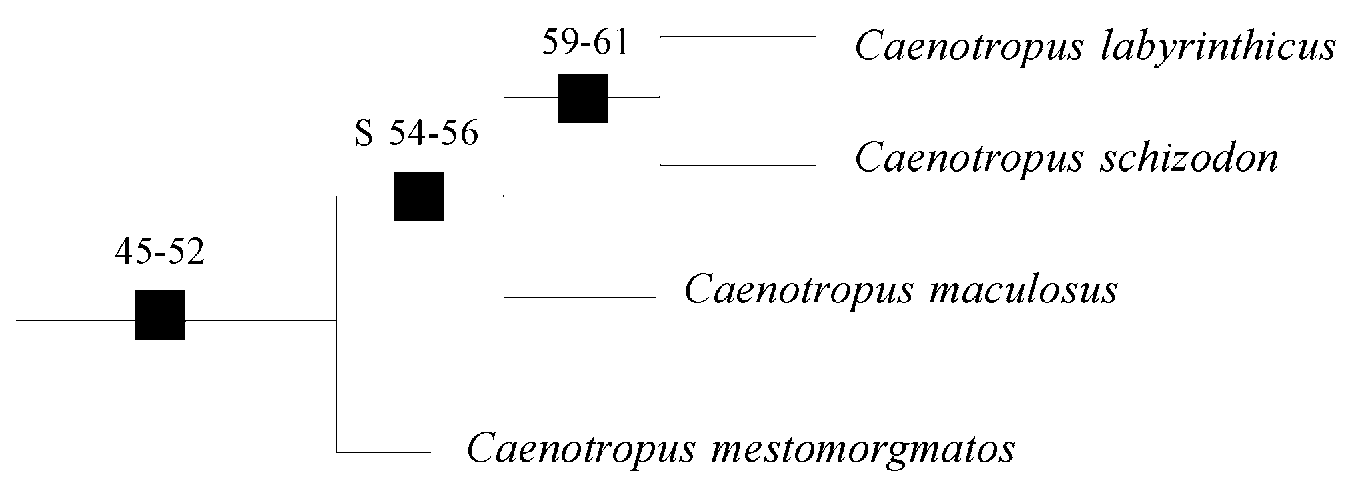
TABLE 1. Morphometrics of Caenotropus schizodon, n. sp.; H = holotype, MCP 29971; n = number of specimens, holotype included; SD = Standard Deviation. * 7 scales in 2 specimens.
| Range | |||
|---|---|---|---|
| Character | n H Low High | Mean | SD |
| Standard Length (mm) | 18 71.5 46.1 90.5 Percents of standard length | 59.0 | 14.766 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Caenotropus schizodon
| Scharcansky, Alexandre & Lucena, Carlos Alberto Santos De 2007 |
Caenotropus labyrinthicus
| Vari 1995: 26 |