
Nuchequula mannusella, CHAKRABARTY & SPARKS, 2007
|
publication ID |
https://doi.org/10.1206/0003-0082(2007)3588[1:PATRON]2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03E06774-4464-403D-FF77-64CF1D02FB51 |
|
treatment provided by |
Carolina |
|
scientific name |
Nuchequula mannusella |
| status |
sp. nov. |
Nuchequula mannusella View in CoL , new species figures 4 View Fig , 5A View Fig , tables 2–3
HOLOTYPE: AMNH 238753 About AMNH , 85.5 mm SL; Taiwan: Chiayi County: Tungshih Fish Market : 23 ° 27 9 1 0 N, 120 ° 08 9 19.3 0 E; TW-6-2006; Prosanta Chakrabarty, Joker K.H. Chiu, John S. Sparks, 22 March 2006.
PARATYPES: AMNH 238754 About AMNH , 15 ex., 72.1– 90.1 mm SL; data as for holotype . AMNH 238755 About AMNH , 16 ex., 62.9–89.7 mm SL; data as for holotype . AMNH 238756 About AMNH , 15 ex., 70.2–94.1 mm SL; data as for holotype . AMNH 238757 About AMNH , 14 ex., 64.8–81.2 mm SL; data as for holotype . AMNH 238758 About AMNH , 20 ex., 77.2–86.9 mm SL; data as for holotype . AMNH 238759 About AMNH , 9 ex., 82.1–95.4 mm SL; data as for holotype . AMNH 238760 About AMNH , 13 ex., 75.1–98.5 mm SL; data as for holotype . AMNH 238761 About AMNH , 1 ex., 71.2 mm SL; Taiwan: Hsinchu City: Motorway 3 north from Taichung, oneand-one-half hours from Taichung, West Coast Hwy: Fishing Harbor in Hsinchu: 24 ° 50 9 55.4 0 N, 120 ° 55 9 13.6 0 E; local fisherman, Prosanta Chakrabarty, Otto Jeng-Di Lee, John S. Sparks, 20 March 2006 . AMNH 238762 About AMNH , 1 ex., 53.4 mm SL; data as for holotype . AMNH 238763 About AMNH , 1 ex., 83.0 mm SL; data as for holotype . AMNH 238764 About AMNH , 1 ex., 88.9 mm SL; data as for holotype . AMNH 238765 About AMNH , 29 ex., 73.6– 100.9 mm SL; data as for holotype . ASIZP 0062322 View Materials , 1 ex., 76.2 mm SL; Taiwan: Fenggang :
TABLE 1 Morphological Transformations Used in the Phylogenetic Analysis of Nuchequula Species (see fig. 3 View Fig )
Pingtung: open sea, at 100 m depth: 22 ° 26 9 N, 120 ° 38 9 E; J.H. Wu, 1 March 2001. ASIZP 0060823 View Materials , 1 ex., 90.9 mm SL; Taiwan: Kaoshiung: Shingda Harbor: open sea: 22 ° 87 9 N, 120 ° 19 9 E; G.J. Xia, 8 June 2000. SIO 06.261 About SIO , 2 ex., 83.7–96.2 mm SL; data as for holotype. (Because much of this collection is from a single market locality, different AMNH numbers were assigned to material to reflect the possibility that different vendors sold fish from diverse sources.) GoogleMaps
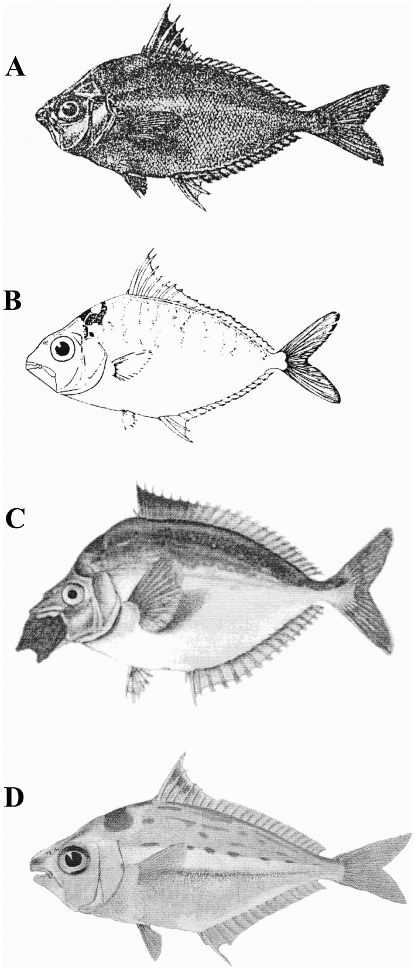
DIAGNOSIS: Nuchequula mannusella is distinguished from its congeners by a conspicuously deep lower jaw with a strongly concave profile that is sharply pointed ventrally (versus a slightly concave or straight lower jaw profile). The black dorsal-fin marking of N. mannusella is uniquely restricted to the anterior edge of the fin, with some scattered melanophores in the dorsal-fin membrane (versus a large blotch over a larger area on the dorsal fin or an absence of markings on the dorsal fin).
DESCRIPTION: Morphometric and meristic data are presented in table 2, and other comparative features are presented in table 3. Nuchequula mannusella is a moderately large (maximum recorded length 101 mm SL), rhomboid-shaped ponyfish. The dorsal and ventral profiles are equally convex. The greatest body depth is reached at the vertical from the dorsal-fin origin to the pelvic-fin origin. There is a concavity above the orbit, formed by the exposed rising dorsal aspect of the supraoccipital crest (i.e., nuchal spine). The dorsal- and pelvic-fin origins are located along the same vertical. The anal-fin origin is located at the vertical from the last ( 5 eighth) dorsalfin spine. The mouth is terminal and directed strongly ventrally when extended. The lower jaw is deep, and the lateral profile is strongly concave. The lips are thin and only somewhat fleshy. The posterior margin of the maxilla is exposed, reaching the vertical through the anterior part of pupil and the dorsal portion of the lower jaw articulation. The exposed part of maxilla is nearly perpendicular with the mouth when retracted. The eyes are large and circular (, 10% SL). The sensory pores around the orbit are moderately well developed; some pores posterior to the orbit reach the preopercle. Sensory canals are also present ventral to the orbit and dorsal to the mouth. Two long sensory canals are present ventral to the nuchal spine. Two closely apposed nasal pores are present anterior to the orbit. The anterior nasal pore is smaller and more rounded than the posterior pore. The preopercular margin is strongly serrated along the ventral margin, with 20–25 spines. The supraorbital ridge bears small spines. There are two short and stout postnasal spines present dorsal to the orbit and posterior to the nasal pores. There are 16 elongate and slender gill rakers present on the lower limb of the first gill arch; the rakers are sparsely denticulated (i.e., bearing few teeth). The longest rakers are about the length of the gill lamellae, whereas the shortest are about half the length of the lamellae. There are 23 total vertebrae (10 precaudal + 13 caudal).
Fins: The dorsal fin has VIII spines and 16 rays. The first dorsal-fin spine is much shorter than the other spines, and 15–20% of the second spine. The length of the second and longest dorsal-fin spine is about 40% of body depth and not conspicuously longer than the third or fourth spine. The second and third spines are more robust, particularly at the base, than the remaining spines. The third and fourth spines possess serrations facilitating interlocking with the preceding spine. The anal fin has III spines and 14 rays. The first anal-fin spine is much shorter than the others, 25–35% of the second. The second anal-fin spine is more robust than the other spines; it is only slightly longer than the third and is about 30% of body depth. The third anal-fin spine possesses serrations facilitating interlocking with the second spine. There is a sheath of asquamate tissue that covers the bases of the spinous portions of the dorsal and anal fins. The pelvic fins are short, and do not reach the origin of the anal fin, and have one associated spine. The caudal fin is deeply forked.
Dentition: The teeth are villiform and arrayed in a narrow band of three to five poorly differentiated rows. The teeth in the lower jaw are slightly recurved, whereas those in the upper jaw are strongly recurved and almost parallel with the roof of the mouth.
Squamation: As for all leiognathids, scales are highly deciduous, making accurate counts and observations problematic. The chest and fins are asquamate. Some specimens possess scales on the cheek and in the interpelvic region. There are scales present in the nuchal region and continuing to the ventrum (not including the chest), which extend posteriorly to the caudal peduncle and onto the caudal fin. The lateral line is complete, extending from the posterior edge of the opercle to the posterior margin of the caudal peduncle, and includes about 55 scales. The pores of the lateral line are well developed. The lateral line is horizontal between the posterodorsal margin of the opercle to slightly beyond the pectoral-fin base. The lateral line is arched slightly from the pectoralfin base posteriorly to the caudal peduncle and is horizontal on the caudal peduncle.
Pigmentation in life: The entire body is silvery (see fig. 5A View Fig ). The leading edge of the
TABLE 2 Morphometric Data for Nuchequula mannusella ( N 5 32)
dorsal fin is black. The spinous region of the dorsal fin is yellow from about three-quarters the length of the second dorsal spine to the distal margin. The entire dorsal fin is yellow distally. The ventral lobe of the caudal fin bears significantly more yellow along its posterior margin than the dorsal lobe. The spinous portion and entire distal margin of the anal fin are yellow. The base of the pectoral fin is yellow. The yellow areas on the fins in some specimens have a reddish hue. The buccal area (mouth, lower jaw) and the region dorsal to the orbit are translucent but sometimes reddish. There is a dark brown or black saddle-shaped, triangular nuchal marking. In some specimens there is a light-blue metallic spangling dorsal to the opercle and a blue ‘‘W’’-shaped marking straddling the dorsal aspect of the lateral line ventral to the spinous portion of the dorsal fin (usually between the fourth and seventh spines). The pectoral-fin axil is black. Dark vertical bands are present on the flanks dorsal to the midline, ranging from broad and nearly straight to vermiculate and moderately curved. These bands sometimes coalesce ventrally.
Pigmentation in preservative: Most specimens comprising the type series have lost their guanine in preservation. Base body coloration is generally tan. In specimens retaining guanine, the body is silvery from slightly above the midline to the ventrum, and the gill covers and suborbital regions are silvery to a vertical through the anterior margin of the orbit. As in life, the dorsal flank markings consist of dark vertical bands that vary from broad and straight to vermiculate and wavy or that form a zigzag pattern. The dorsal markings are most prominent above the lateral line. A dark blotch is present above the upper lip anterior to the nasal pores. The pectoral-fin axil is black or dark brown. Scattered melanophores are present ventrally on the body; they are obscured when guanine is retained. A pigment-free, mitten-shaped region is visible on specimens lacking guanine posterior to the pectoral-fin base and between the pelvic- and anal-fin origins. The pigment-free region is not
TABLE 3 Comparative Features of Species of Nuchequula Based on Type Material and Original Descriptions translucent but is conspicuous because it is the only section ventral to the midline that lacks melanophores. Black pigmentation is present on the leading edge of the dorsal fin, and scattered melanophores are present on the dorsal-fin membrane. A conspicuous black or dark brown triangular (sometimes rounded ventrally) marking is present in the nuchal area. The nuchal mark usually reaches slightly beyond the posterior margin of the nuchal spine and to about the midpoint between the nuchal spine and the lateral line ventrally.
DISTRIBUTION: Currently known only from the coastal waters of Taiwan ( Hsinchu, Pingtung, Kaoshiung, and from market collections at Tungshih).
ETYMOLOGY: From the Latin mannus (meaning ‘‘pony’’) in reference to the common name for the family, and sella (meaning ‘‘saddle’’), describing the shape of the nuchal marking.
REMARKS: One paratype, AMNH 238761, was collected by a local fisherman using hook and line and observed alive. One of the unusual aspects of the observation was an audible chirping sound made by the fish. Although little can be made of this short observation, it may be important for future reference. Visual communication through bioluminescent flashing is well documented in ponyfishes ( Woodland et al., 2002; Sasaki et al., 2003), and the possibility of an auditory component in ponyfish communication would also be noteworthy. Notably, the common name of this fish in Singapore and other parts of the Malay Peninsula is ‘‘ kekek ’’ the onomatopoeic moniker linked to the chirping sound.
Nuchequula blochii (Valenciennes, in Cuvier & Valenciennes, 1835) figures 6A View Fig , 7 View Fig , tables 3–4
Equula blochii: Valenciennes View in CoL , in Cuvier and Valenciennes, 1835: 84; Day, 1865: 105; Day, 1875: 238, 241, plate LII, fig. 5 View Fig .
Eubleekeria (Nuchequula) blochii: Whitley, 1932: 109 .
DIAGNOSIS: Distinguished from its congeners, except Nuchequula pan , by the presence of scales on the chest. Nuchequula blochii is distinguished from N. pan by the absence of longitudinal lines on the flank.
DESCRIPTION: Comparative features are presented in table 3; morphometric and meristic data are presented in table 4. Nuchequula blochii was described as having an oblong body, a concave lower jaw, and strong dorsal- and anal-fin spines. (We base all elements of this description exclusively on our examination of part of the syntypic series for this species.) The mouth is terminal and directed ventrally when extended. The teeth are villiform and arrayed in a narrow band. Body depth of the syntypes is about 40% of SL. The length of the second dorsal-fin spine is 50% of body depth, and the length of the second anal-fin spine is 46% of body depth (intact in only one specimen). The length of the first dorsal-fin spine is about 15% of the second spine. The length of the first anal-fin spine is 25% of the second. The dorsal- and anal-fin spines are robust (the second and third more so than the others). In lateral view, a slight concavity on the head is present above the orbit. The dorsal and ventral profiles are about equally convex. The caudal fin is deeply forked. The depth of the caudal peduncle is about 6% of SL. The lateral line is complete from the opercle to the posterior margin of the caudal peduncle and parallel with the shape of the back. The lips are thin and not fleshy. The dorsal- and pelvic-fin origins are located along the same vertical. The anal-fin origin is located on the vertical from the eighth dorsal-fin spine. There are 16 rakers on the lower limb of the first gill arch. These rakers are about one-half to three-quarters the length of the gill lamellae. Some scales are scattered along the body (most are apparently lost due to preservation), and scales are still present on the chest and nape. The lateral profile of the lower jaw is concave. Two robust postnasal spines are present dorsal to the orbit and posterior to the nasal pores. The supraorbital ridge is weakly serrated. The width of the body ranges from about 8–12% of SL.
Pigmentation pattern in life: We lack fresh material for study and quote this description from Day (1875), who examined specimens from Malabar, the type locality: ‘‘silvery, with a dark brown blotch over nape, and a black mark in the upper half of the spinous dorsal fin from the third to the sixth spines. Vertical zig-zag yellow lines on the back and sides, which fade soon after death: base of pectoral posteriorly dark coloured.’’
Pigmentation in preservative: Coloration of N. blochii was originally described as being similar to that of most other ponyfish species,
TABLE 4 Comparative Morphometric and Meristic Data for Nuchequula Species Based on Descriptions in the Literature (marked with asterisk, *) or Our Measurement of Types or Topotypic Material with gray or lead coloration on the back (Valenciennes, in Cuvier and Valenciennes, 1835). A bright silver longitudinal band was described as separating the gray dorsal and silvery ventral areas. Valenciennes (in Cuvier and Valenciennes, 1835) noted that this species possesses a black blotch on the dorsal fin, but he did not comment on the presence of a nuchal spot. Although the two type specimens we have available (MNHN A-6757, 71.4 mm SL; MNHN A6759, 67.7 mm SL) are quite faded in preservative, they retain a dark brownish marking below the nuchal spine. The two syntypes are covered in guanine ventral to the midline, with some yellowish scales present on the chest and along the flank. The fins are a faint opaque yellow. No remnant of a blotch on the dorsal fin remains in either specimen.
DISTRIBUTION: From western India to Myanmar; Day (1875) had specimens from ‘‘ Bombay, Malabar, Madras, Calcutta, and Akyab, and many young from the Sunderbunds [Sunderbans].’’
REMARKS: Day (1875) confirmed the presence of a nuchal blotch in N. blochii , commenting that the species is ‘‘silvery with a dark brown blotch over the nape.’’ Day (1875) does not mention whether he examined the syntypes of this species, but he does mention that he looked at specimens from Malabar, the type locality. Given that Day (1875) based his redescription of this species on topotypic material and that the features he presents are in agreement with both the original description of Valenciennes and the type series, we conclude that Day (1875) and Valenciennes (in Cuvier and Valenciennes, 1835) were describing the same species. The additional features that Day presents are most likely a consequence of examining fresher material than that available to Cuvier and Valenciennes at the time of their original description of N. blochii .
We initially identified individuals of the new species described here, Nuchequula mannusella , as N. blochii in the field. Only when we examined the type material and the original description of N. blochii did we recognize that our Taiwan material represented a novel species.
Another potentially novel species is what has been identified as Leiognathus blochii in Australia. Jones (1985) notes that her description of N. blochii from Australia ‘‘differs in some respects from accounts of L. blochii in the literature. Day (1875) and James (1975), for instance, describe L. blochii as having a distinct black blotch on the spinous dorsal fin, whereas in the Australian specimens the dorsal fin is colourless except for a yellow streak at midheight.’’ Jones appears to have been unaware that in the original description of Nuchequula blochii, Valenciennes provided one of the few diagnostic characters useful for distinguishing N. blochii : the dorsal-fin pigmentation. It is unclear from Jones’s (1985) description exactly which species she is referring to, and we have been unable to obtain any of the specimens she examined to corroborate our hypothesis. The main feature she uses to distinguish her Australian ‘‘ L. blochii ’’ from N. decora is the possession of scales on the chest in the former, the same feature used by Day (1875) to distinguish N. blochii from what he referred to as Leiognathus brevirostris .
We have observed that a mitten-shaped melanophore-free region is present posteroventrally from the pectoral-fin base in all species of Nuchequula except N. blochii . This pigment-free area can be observed only in specimens that lack guanine. Unfortunately, all specimens of N. blochii that we have for observation (which we restrict to syntypes) have guanine that obscures this ventral melanophore pattern (assuming that it is present in N. blochii ). This feature is a synapomorphy of Nuchequula , as it has not been observed in other leiognathid species. We note, however, that other ponyfish species have been observed with differently shaped melanophore-free regions, which may prove useful for resolving relationships within the group.
Nuchequula decora ( De Vis, 1884) View in CoL figures 6B View Fig , 8 View Fig , tables 3–4
Equula decora: De Vis, 1884: 543 View in CoL .
Leiognathus decorus: Jones, 1985: 579–581 View in CoL , fig. 8. View Fig
DIAGNOSIS: Nuchequula decora is distinguished from congeners by a nuchal spot that is U-shaped or irregularly shaped and concentrated (versus diffuse and rounded or triangular). A central (occasionally more anterior) portion of the nuchal spine associat- ed with the nuchal spot is pigment-free, unlike other Nuchequula species , in which it is fully pigmented. In addition, N. decora is the only member of Nuchequula lacking dorsal-fin markings.
DESCRIPTION: Comparative features are presented in table 3; morphometric and meristic data are presented in table 4. The original description of Nuchequula decora by De Vis (1884) is not informative in terms of distinguishing the species from other leiognathids in that no diagnostic features are presented. Jones (1985), who examined type material and fresh material from the same general region (northern Australian waters) of the type series, describes N. decora as having ‘‘body depth between 43–55%, chest and suborbital naked … second dorsal spine approx. half body depth … gill rakers slightly less than half the length of gill lamellae.’’ Our examination of material identified by Jones as ‘‘ Leiognathus decorus ’’ revealed that the mouth is terminal and directed ventrally when extended. (We base our redescription exclusively on material identified by Jones; see Remarks for a justification.) Teeth are villiform and in a narrow band of three to four poorly differentiated rows. Body depth ranges between 45% and 55% of SL. The length of the second dorsal-fin spine is 35–50% of the body depth; the length of the second anal-fin spine is about 30–40% of body depth. The length of the first dorsal-fin spine is about 12–15% of the second spine. The length of the first analfin spine is about 10–20% of the second. The dorsal- and anal-fin spines are weak and often are broken at the tips. In lateral view, a weak concavity is present on the head dorsal to the orbit. The caudal fin is deeply forked. Depth of the caudal peduncle is about 6–7% of SL. The lateral line is complete from the opercle to the posterior margin of the caudal peduncle and is parallel with the shape of the back. The lips are thin but somewhat fleshy. The dorsal- and pelvic-fin origins are located along the same vertical. The anal-fin origin is located on a vertical between the seventh and eighth dorsal-fin spines. There are 16 rakers on the lower limb of the first gill arch. These rakers are about one-quarter to one-half the length of the gill lamellae. Scales are absent on the chest and head but are present throughout much of the rest of the body, including the nape and caudal peduncle. The profile of the lower jaw in lateral view is slightly concave to straight. Two small postnasal spines are present dorsal to the orbit and posterior to the nasal pores; these spines do not protrude much from the body. The supraorbital ridge is very weakly serrated and is not rough to the touch.
Pigmentation pattern in life: We lack fresh material for examination and defer to Jones (1985) for a description of live coloration. Jones (1985) describes Nuchequula decora with ‘‘body silver, upper half with irregular dark brown wavy to zig-zag vertical lines; brown blotch across nape... outer half of spinous dorsal and anal fins yellow, colour continued along margins of rays, caudal fin with dusky yellow margin; other fins colourless.’’
Pigmentation in preservative: The body is silvery with guanine that becomes more concentrated ventral to the midline. Above the midline, the body is generally light brown. A U-shaped nuchal marking, or a nuchal spot with a small pigment-free region straddling the nuchal spine at its midpoint (or occasionally more anteriorly), is characteristic of this species. Pigmentation pattern on the dorsal flank consists of dark, wavy horizontal lines that are more strongly curved and more concentrated dorsal to the lateral line than ventrally. There is a concentration of melanophores dorsal to the upper lip and anterior to the orbit, as well as in the pectoral-fin axil. The dorsal fin in most preserved specimens is pigment-free, but in some specimens there is a light scattering of melanophores in the membranes of the fin and on the anterior edge of the second dorsal-fin spine.
DISTRIBUTION: Coastal waters surrounding northern Australia. Jones (1985) illustrates the Australian distribution as extending from north of Exmouth in Western Australia to north of Brisbane in Queensland.
REMARKS: In discussing the pigmentation pattern of Nuchequula decora, De Vis (1884) mentions ‘‘[t]hree dark lines, terminating very obscure bands, across the back, at the base of the soft dorsal and one on the edge of the caudal peduncle.’’ It appears that De Vis was referring to the distinctive vertical bands on the upper flanks that are characteristic of this species. The fact that he mentions only four lines may indicate that he did not have access to fresh material (see Jones, 1985; Whitley, 1932), as these markings fade in preservation. The type series, a lectotype (QM I.1698) and two paralectotypes (QM I.4877), comprises all small specimens ( 24–40 mm SL; from fig. 8 View Fig , Jones, 1985), and individuals of N. decora of this size range often lack the characteristic pigmentation pattern of the adults, even as fresh material (Jeff Johnson, QM, personal commun.).
Jones (1985) states in reference to the original description of Equula decora by De Vis (1884), ‘‘Although no mention is made of a nuchal blotch, this pigmentation often fades on preservation, leaving only the more persistent vertical bands across the back. The body proportions of the type-specimens are consistent with the species as described above.’’ Half of the original six specimens examined by De Vis are lost. The remaining three specimens, all of which are small ( 24–40 mm SL; from Jones, 1985: fig. 8 View Fig ), have faded in preservative and lack pigmentation. Whitley (1932) commented, ‘‘[T]he scales are now rubbed off the types of Equula decora and it is impossible to observe whether the breast was naked or scaly … [T]he colours have now faded.’’ Jones (1985) utilizes a number of measurements and other features to tie the type series and original description of De Vis to what she recognized as Nuchequula decora . We consider Jones’s (1985) redescription as an accurate depiction of Nuchequula decora that is in agreement with the original description of De Vis (1884). Jones examined the type series as well as fresh material from the region of the type locality, which lends credibility to her redescription.





| AMNH |
American Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nuchequula mannusella
| CHAKRABARTY, PROSANTA & SPARKS, JOHN S. 2007 |
Leiognathus decorus : Jones, 1985: 579–581
| Jones, G. 1985: 581 |
Eubleekeria (Nuchequula) blochii :
| Whitley, G. P. 1932: 109 |
Equula decora : De Vis, 1884: 543
| De Vis, C. W. 1884: 543 |
Equula blochii :
| Day, F. 1875: 238 |
| Day, F. 1865: 105 |
| Valenciennes, A. 1835: 84 |