
Necromantis fragmentum Ravel, 2016
|
publication ID |
https://doi.org/10.5252/g2016n3a3 |
|
publication LSID |
urn:lsid:zoobank.org:pub:FC07ACBE-03F7-414A-BB64-1BB0711766BF |
|
persistent identifier |
https://treatment.plazi.org/id/03E07259-FF88-FFBA-FC4B-F955CE7FFC07 |
|
treatment provided by |
Felipe |
|
scientific name |
Necromantis fragmentum Ravel |
| status |
sp. nov. |
? Necromantis fragmentum Ravel , n. sp.
( Figs 4 View FIG , 5 View FIG ; Tableau 1)
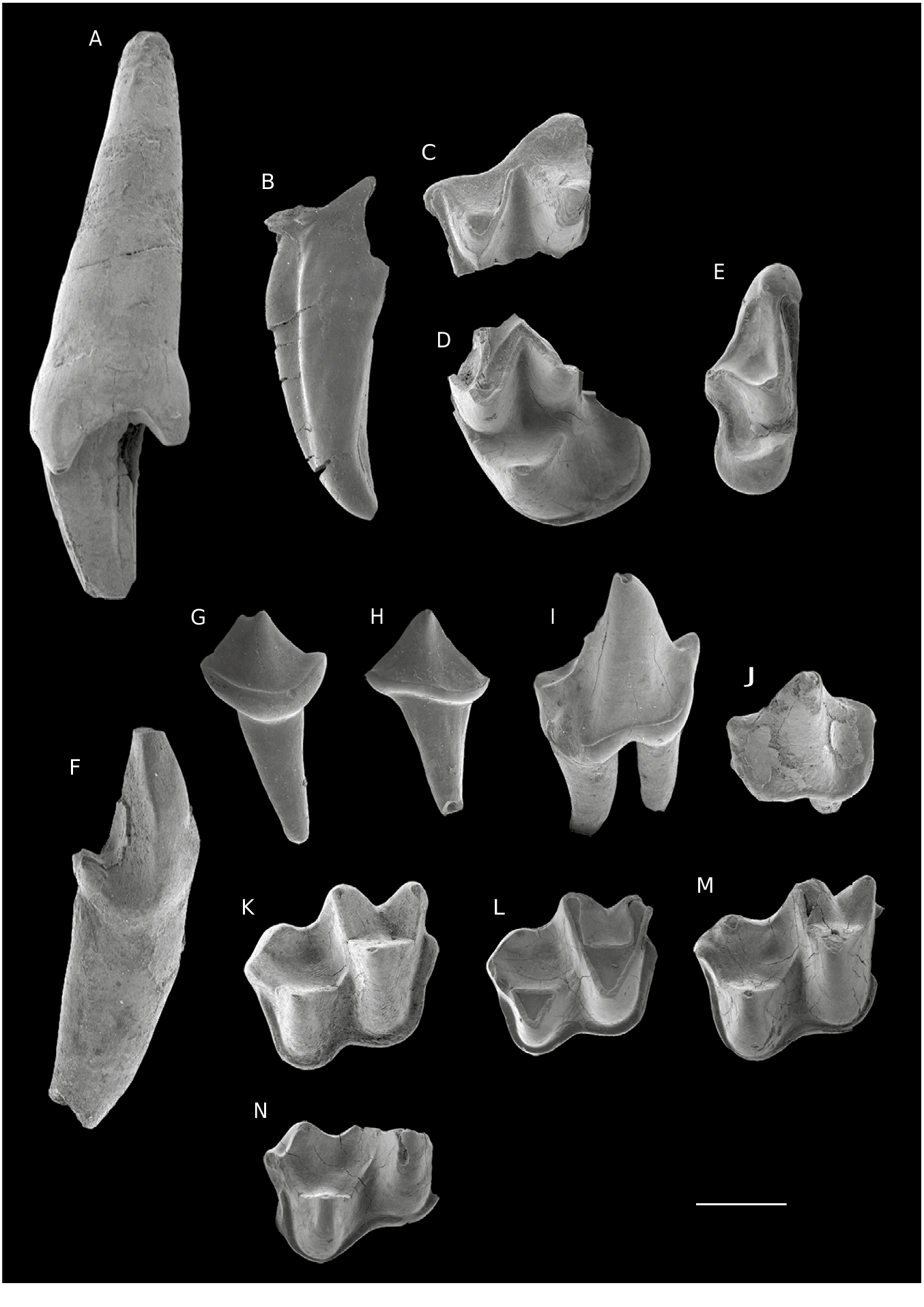
HOLOTYPE. — En l’absence de molaire complète, l’holotype considéré ici est la canine supérieure isolée droite CBI-2-010 marquant l’originalité de l’espèce nouvellement créée ( Figs 4A View FIG ; 5A View FIG ).
MATÉRIEL EXAMINÉ. — CBI-1-243 (tubercule de C1 gauche; Figs 4B View FIG ; 5 B View FIG ); CBI-2-007 (ectolophe de M1/2 gauche; Figs 4C View FIG ; 5C View FIG ); CBI-2-008
Tunisie. Abréviations: L, longueur; l, largeur; H, hauteur; Ltdr, longueur du trigonide; ltdr, largeur du trigonide; Ltdl, longueur du talonide; ltdl, largeur du talonide. Le signe * derrière une valeur indique un biais dans la mesure causé par l’altération de la couronne.
(partie linguale d’une M1/2 gauche; Figs 4D View FIG ; 5 D View FIG ); CBI-2-009 (M3 droite; Figs 4E View FIG ; 5 E View FIG ); CBI-2-016 (c1 droite; Figs 4K View FIG ; 5 F View FIG ); CBI-2-013 (p4 droite; Figs 4 I View FIG ; 5I View FIG ); CBI-2-014 (p4 gauche; Fig.5J View FIG ); CBI-2-017 ( Figs 4J View FIG ; 5 G View FIG ), CBI-2-018 ( Fig.5H View FIG ), CBI-2-019,CBI-2-020 et CBI-2- 022 (p2s gauches); CBI-2-021 et CBI-2-023 (p2s droites), CBI-1-244 ( Figs 4F View FIG ; 5 K View FIG ) et CBI-3-003 (m1s droites Figs 4G View FIG ; 5 L View FIG ); CBI-2-011 (m2 droite; Figs 4 H View FIG ; 5 M View FIG ); CBI-2-012 (m1/2 droite; Fig. 5N View FIG ).
ÉTYMOLOGIE. — Du latin « fragmentum » qui veut dire fragment ou débris, faisant référence au caractère fragmentaire du matériel attribué à ce taxon.
LOCALITÉ TYPE ET ÂGE. — Chambi (CBI) loci 1, 2 et 3, Éocène inférieur terminal-Éocène moyen basal, Djebel Chambi, situé dans la région de Kasserine, Tunisie.
DIAGNOSE. — Chiroptère avec une denture robuste, canine supérieure munie de deux tubercules secondaires puissants, une molaire supérieure avec un léger ectoflexus et un bord buccal très incliné mésiolingualement, une p4 présentant une ébauche de paraconide et un bassin du talonide plus développé.
DESCRIPTION
Ce taxon est représenté par les plus grands spécimens provenant des loci 2 et 3 de Chambi, et le deuxième plus grand du locus 1 après l’espèce Witwatia sigei Ravel, Marivaux, Tabuce, Ben Haj Ali, Essid & Vianey-Liaud, 2012 ( Ravel et al. 2012, 2015; Tableau 1).
Les canines supérieures CBI-2-010 et CBI-1-243 ( Figs 4A, B View FIG ; 5A, B View FIG ) sont attribuées à cette espèce en raison de leur taille importante et leur robustesse, compatibles avec les dents jugales associées. CBI-2-010 est très massive. La couronne est composée de trois tubercules: un principal en position mésiobuccale et deux secondaires assez forts situés aux bases mésio- et distolinguale du tubercule principal. Le tubercule principal est muni de deux faces: une face buccale convexe s’opposant à une face linguale quasi plate. Mésialement à la face buccale, un large sillon est visible. Le cingulum épais fait la liaison entre les deux cuspides secondaires lingualement. Il est plus fin et discontinu dans sa partie buccale. CBI-1-243 est une partie apicale du tubercule principal d’une canine ( Figs 4B View FIG ; 5 B View FIG ). Le tubercule est modérément recourbé vers la partie distale au niveau de l’apex. La face mésiobuccale est bombée, et porte un sillon s’étalant sur les trois quarts de la hauteur du tubercule comme sur CBI-2- 010 ( Figs 4A View FIG ; 5 A View FIG ). La face linguale, plate, est délimitée par une crête mésiale et une crête distale tranchantes.
CBI-2-007 est un fragment de dent correspondant à un ectolophe ( Figs 4C View FIG ; 5C View FIG ). Son bord buccal est très incliné mésiolingualement au niveau du métacône puis s’aplanit au niveau du paracône. Le parastyle forme un «crochet » très recourbé vers l’extrémité mésiale. Il se positionne plus lingualement que le mésostyle et le métastyle. Le mésostyle, en partie effacé par l’usure de l’émail, est peu développé et en retrait par rapport au bord buccal. Le métastyle, en partie cassé, continue buccalement la postmétacrête de façon rectiligne. L’ectolophe forme un « W » légèrement comprimé dans sa partie distale («V » mésial plus ouvert que le « V » distal). Le paracône se positionne plus lingualement et apparaît plus comprimé buccolingualement par rapport au métacône. Les deux tubercules buccaux sont espacés par une longue et large protofosse qui s’étend jusqu’à la base du mésostyle. Le précingulum est modérément large et s’oppose au postcingulum qui est beaucoup plus étroit.
Le spécimen CBI-2-008 est une moitié linguale de M1/2, dont une partie de la région du mésostyle et les flancs linguaux du paracône et du métacône ont été conservés ( Figs 4D View FIG ; 5 D View FIG ). Comme il a été observé sur l’ectolophe CBI-2-007, le paracône est plus lingual que le métacône. Les deux tubercules sont néanmoins plus rapprochés limitant l’extension longitudinale de la protofosse. Le mésostyle forme un angle obtus en retrait du bord buccal. Le protocône est droit, haut et très pointu. La préprotocrête rectiligne remonte buccalement depuis le sommet du protocône pour rejoindre le mince précingulum. La postprotocrête s’oriente distobuccalement de façon symétrique à la préprotocrête. Elle se termine lingualement par rapport à la base du métacône laissant ainsi une ouverture distale à la protofosse. Le bassin du talon se projette distalement. Il s’étend vers la partie buccale jusqu’à la terminaison linguale du postcingulum. Le bassin du talon est ceinturé par un large cingulum lingual. Ce cingulum effectue un renflement distolingual au protocône sans pour autant réaliser un tubercule bien individualisé.
La M3, CBI-2-009 ( Figs 4E View FIG ; 5 E View FIG ), est complète est bien préservée. Cette dernière molaire supérieure apparaît plus réduite en largeur et en longueur que les deux autres spécimens. Son développement est néanmoins à tendance transversale. Le bord buccal de la dent est très incliné distolingualement et est muni d’un fort cingulum buccal continu. Le parastyle est particulièrement bien développé et prend la forme d’un large crochet projeté buccomésialement. Le paracône est réduit et déplacé vers le bord mésial. L’ectolophe, dépourvu de métacône, se termine distalement par un fragment de la prémétacrête. Le mésostyle conservé est peu développé et légèrement en retrait par rapport au bord buccal. Le protocône est faible et déjeté mésialement. Il se positionne ainsi sur le même axe buccolingual que le paracône. La préprotocrête se connecte mésiobuccalement au précingulum tandis que la postprotocrête se courbe distobuccalement pour rejoindre le postcingulum. Ce dernier, assez court, débute buccalement à la base de l’extrémité distale de la prémétacrête. La bordure linguale de la M3 est arrondie et dépourvue de cingulum.
La c1 (CBI-2-016; Figs 4K View FIG ; 5 F View FIG ) possède un unique tubercule droit décomposé en une face mésiobuccale convexe et une face distolinguale plane. Les deux faces sont délimitées par une crête mésiale et une crête distale qui s’étendent depuis l’apex du tubercule. Le cingulum visible sur tout le pourtour de la couronne forme un bourrelet distolingual qui délimite un bassin étroit situé à la base de la cuspide.
CBI-2-017 à CBI-2-018 sont des dents possédant des couronnes très simples, unicuspidées, et dont la morphologie rappelle celle de p2 ( Fig. 5G, H View FIG ). Ces prémolaires ont une couronne ovale en vue occlusale. La cuspide se décompose en deux faces (une face buccale convexe s’opposant à une face linguale sub-plate) délimitées mésialement et distalement par des crêtes sagittales. La crête mésiale est légèrement plus courte et tranchante que la crête distale. La couronne est cernée par un épais cingulide continu, courbé buccalement et rectiligne lingualement.
Les p4s identifiées (CBI-2-013 et CBI-2-014; Figs 4I View FIG ; 5 I, J View FIG ) sont biradiculées et possèdent une couronne allongée mésiodistalement (cependant moins longue que les m1-2s) et très étroite transversalement. La cuspide principale (i.e., protoconide) est haute et robuste. En position mésiale, le paraconide est légèrement plus lingual que le protoconide. Il s’élève au quart de la hauteur du protoconide. Les deux cuspides (protoconide et paraconide) sont connectées par une crête ayant une orientation à prédominance mésiale depuis l’apex du protoconide. À l’opposé, une large crête s’étend distolingualement depuis la pointe du protoconide. Elle effectue un renflement à mi-hauteur de la cuspide principale (comme un vestige de métaconide). La partie distale de la couronne est composée d’un talonide vestigial large et très projeté distalement. Le talonide se résume en un bassin traversé par une courte crête qui s’étend mésiodistalement depuis la base du protoconide jusqu’à l’extrémité distale de la couronne. Le bassin est circonscrit par un large cingulide qui s’étend jusqu’à la base mésiobuccale du protoconide.
Le trigonide des m1-2s est très massif et domine le talonide ( Figs 4 View FIG ; 5 View FIG ). Le paraconide est de taille quasi équivalente au métaconide. La projection mésiale du paraconide est plus prononcée sur m1 que sur m2, formant ainsi un trigonide plus ouvert sur cette dernière. Ceci est l’unique différence significative entre les deux types de molaires. Le protoconide est la cuspide dominante des molaires inférieures en termes de hauteur et de volume. Le talonide est de largeur et longueur comparables au trigonide. L’hypoconide se situe sur le même axe mésiodistal que le protoconide. La forte différence de hauteur entre l’hypoconide et le protoconide est associée à une flexion très prononcée du bord buccal de la couronne. La crête oblique, relativement courte, remonte depuis l’hypoconide jusqu’au niveau de la jonction entre la métacristide et la postprotocristide. L’entoconide, peu développé, se situe sur le même axe linguobuccal que l’hypoconide. L’hypoconulide, légèrement plus réduit que l’entoconide, occupe une position médiane et très distale par rapport à l’entoconide et à l’hypoconide. Il est relié à l’hypoconide et à l’entoconide par la postcristide. La position de l’hypoconulide et l’extension de la postcristide est caractéristique d’une structure nécromantodonte. La position de l’hypoconulide est plus linguale sur CBI-2-011 et CBI-2-012, soulignant ainsi une certaine variabilité de la structure du talonide au sein de l’espèce. Sur CBI-2-011, la crête reliant l’hypoconulide et l’entoconide n’apparaît pas, rappelant ainsi un aspect plus nyctalodonte. Le cingulide est continu et épais sur toutes les bordures mésiale, buccale et distale de la couronne .
COMPARAISON ET DISCUSSION
L’aspect robuste des dents de? Necromantis fragmentum Ravel , n. sp. ainsi que la présence d’une p4 non molariforme sont des caractéristiques compatibles avec la diagnose des Necromantidae proposée par Sigé (2011a). La famille regroupe les genres Cryptobune Sigé, 2011 et Necromantis sur la base de caractères soulignant un aspect carnassier. Cryptobune est documenté par un seul spécimen de très grande taille (un dentaire préservant p4 et m1-3) provenant d’une localité inconnue du Quercy ( Sigé 2011a). L’étude de la morphologie dentaire de ce spécimen a conduit Sigé (2011a) à créer la famille des Necromantidae incluant également le genre Necromantis .? Necromantis fragmentum Ravel , n. sp. de Chambi s’écarte de Cryptobune par sa taille beaucoup plus réduite, le trigonide moins compressé buccolingualement, des cuspides mieux individualisées et hautes, et le développement plus important du talonide des molaires inférieures et de p4.
Le genre Necromantis (espèce type N. adichaster ) fut créé par Weithofer en 1887 sur la base d’un matériel très fragmentaire provenant de différentes poches karstiques datées du Paléogène du Quercy. Les spécimens représentatifs de ce taxon sont les plus grands parmi les faunes de chiroptères très diversifiées du Quercy. L’étude d’un nouveau matériel issu des anciennes et nouvelles collections ont permis d’identifier le genre Necromantis depuis l’Éocène moyen (Cuzal, MP13) jusqu’à l’Éocène supérieur (Rosière 5, MP 17b) regroupant plusieurs espèces ( N. adichaster , N. marandati et N. gezei ; Weithofer 1887; Hand et al. 2012; Maitre 2014). Malgré le caractère très fragmentaire du taxon du Djebel Chambi, il est possible d’identifier de nombreuses affinités avec le genre Necromantis européen:
– C1 massive et recourbée distalement;
– un seul ectoflexus;
– mesostyle peu développé et légèrement en retrait du bord buccal;
– bord buccal nettement moins long que le bord lingual; – postprotocrête relativement courte;
– protofosse ouverte distalement (semblable à N. adichaster , les deux autres espèces ont une protofosse complètement fermée);
– extension du bassin du talon de la molaire supérieure; – cingulum lingual arrondi et non cuspidé;
– M3 transverse;
– ectolophe de M3 dépourvu de métacône et de métacrête; – protocône réduit de M3 et projeté mésialement (positionné sur le même axe buccolingual que le métacône);
– p4 allongée et étroite;
– trigonide massif et plus haut que le talonide, mais de longueur et de largeur équivalentes à celles du talonide; – paraconide et métaconide de taille similaire;
– entoconide nettement inférieur au métaconide;
– hypoconide moins volumineux que le protoconide et légèrement plus buccal;
– molaires inférieures nécromantodontes.
Cependant les fossiles tunisiens présentent également des différences marquées en comparaison des formes européennes: – C1 muni de deux fortes cuspides secondaires;
– bord buccal incliné mésiolingualement (cependant assez semblable au spécimen PRRR-70 du Quercy; Maitre 2014: pl. 25f et Hand et al. 2012: fig. 6.10D);
– ectoflexus nettement moins prononcé;
– cingulum lingual s’étendant plus mésialement à la base du protocône;
– M3 relativement plus longue;
– présence d’un paraconide sur p4;
– bassin du talonide de p4 plus développé;
– talonide légèrement plus étendu.
L’absence de rangées dentaires complètes ne permet pas d’aller plus en avant dans la comparaison. Les nombreuses similitudes observées entre les spécimens fossiles de Tunisie et Necromantis suggèrent néanmoins une étroite affiliation entre ces taxons et leur appartenance au même genre. Les divergences morphologiques moins nombreuses soulignent toutefois l’originalité du taxon tunisien notamment au niveau de la C1 qui porte deux cuspides secondaires. Le talonide légèrement plus développé et la présence d’un métaconide sur la p4 confèrent à? N. fragmentum Ravel , n. sp. un aspect archaïque. Maitre (2014) a identifié deux lignées parmi cet ensemble générique. Ainsi la spéciation de N. adichaster à partir de N. marandati se traduit par un raccourcissement de la postprotocrête et une ouverture distale de la protofosse. Cette hypothèse est remise en question par le spécimen CBI-2-008, qui malgré son ancienneté vis-à-vis des deux taxons susnommés présente également une protofosse ouverte distalement.
Le genre Necromantis fut tout d’abord rapproché des Phyllostomidae Gray, 1825 et plus précisément des Phyllostominae (par ex.: Trachops fuliginosus Gray, 1847 , Chrotopterus auritus Peters, 1856 ) par Weithofer (1887) sur les bases de la présence d’une p3 résiduelle déjetée vers le bord lingual, d’une réduction du talonide des m2s et m3s, et de la réduction du nombre d’incisives inférieures (au nombre de deux sur chaque mandibule). Cependant comme le souligne Winge (1893) et Leche (1911), ces caractères sont également partagés avec d’autres familles actuelles (par ex.: Hipposideridae , Rhinolophidae , Megadermatidae ).? Necromantis fragmentum Ravel , n. sp. diffère des Phylostominae en ayant:
– une canine supérieure tri-tuberculée;
– un ectoflexus simple et peu prononcé;
– une forte inclinaison du bord buccal de la molaire supérieure;
– un mésostyle en retrait du bord buccal;
– une p4 allongée pourvue d’un métaconide;
– un cingulum lingual plus étendu mésialement;
– une courte postprotocrête ne se connectant à aucune autre structure.
Ces différences très marquées s’ajoutant à celle que l’on trouve entre les autres espèces mieux documentées de Necromantis et les Phyllostomidae , écartent toute affiliation possible entre le fossile tunisien et le groupe sud-américain actuel. Sur la base d’une argumentation plus poussée et d’un matériel plus fourni, Revilliod (1920) a montré les similitudes entre Necromantis et les Megadermatidae , et plus particulièrement l’espèce Macroderma gigas .? Necromantis fragmentum Ravel , n. sp., comme les autres espèces du même genre, partage avec les Megadermatidae : un mésostyle en retrait du bord buccal, la protofosse étroite mais ouverte distalement, un talon très projeté distalement constitué d’un large bassin, une M3 réduite et étirée transversalement, une p4 allongée, un trigonide puissant et une forte réduction de l’entoconide. En plus de ces similitudes observées, la canine CBI-2-010 attribuée à? N. fragmentum Ravel , n. sp. a la particularité de posséder deux tubercules secondaires.
, Cette configuration est partagée avec les Megadermatidae qui possèdent également deux tubercules secondaires (un très fort en position distale du tubercule principal et le second en position mésiolinguale, formé par le cingulum). L’attribution au genre Necromantis de la canine supérieure de Chambi, si elle venait à être confirmée, indiquerait que Necromantis et les Megadermatidae partagent un ancêtre commun. Toutefois, Necromantis ne possèdent pas les caractéristiques dentaires qui définissent le groupe des Megadermatidae , qui sont:
– ectolophe ayant une forte action tranchante: le paracône et le métacône sont peu développés et sont très rapprochés l’un de l’autre;
– postmétacrête deux fois plus longue que la préparacrête et projetée distolingualement(fonction spécialisée de cisaillement); – protocône très réduit;
– postcingulum absent;
– P4 dépourvu de cuspides secondaires;
– talonide des molaires inférieures réduit par rapport au trigonide; – trigonide de m1 très ouvert avec un paraconide très déjeté mésialement.
De plus l’espèce provenant de Chambi s’écarte des Megadermatidae par:
– le tubercule mésiolingual de C1 plus développé;
– le tubercule distolingual de C1 déplacé lingualement (il est dans l’axe du tubercule principal sur les canines de Megadermatidae );
– le bord buccal fortement incliné mésiolingualement;
– le mésostyle moins développé.
Les différences énoncées ci-dessus s’ajoutent à celles relevées sur les crânes par Hand et al. (2012) entre les Mégadermatidae et Necromantis adichaster . L’accumulation de ces divergences ne permet pas d’inclure le genre Necromantis dans la famille des Megadermatidae ,et conforte le statut familial des Necromantidae .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Necromantis fragmentum Ravel
| Ravel, Anthony, Adaci, Mohammed, Bensalah, Mustapha, Charruault, Anne-Lise, Essid, El Mabrouk, Ammar, Hayet Khayati, Marzougui, Wissem, Mahboubi, Mohammed, Mebrouk, Fateh, Merzeraud, Gilles, Vianey-Liaud, Monique, Tabuce, Rodolphe & Marivaux, Laurent 2016 |
HIPPOSIDERIDAE
| Miller 1907 |