
Trimma meranyx, Winterbottom, Richard, Erdmann, Mark V. & Dita Cahyani, N. K., 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3838.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:5F2E661D-1D8B-4574-8008-70BD0E163734 |
|
DOI |
https://doi.org/10.5281/zenodo.5693548 |
|
persistent identifier |
https://treatment.plazi.org/id/21727606-B099-43D7-B605-E586F864BA35 |
|
taxon LSID |
lsid:zoobank.org:act:21727606-B099-43D7-B605-E586F864BA35 |
|
treatment provided by |
Plazi |
|
scientific name |
Trimma meranyx |
| status |
sp. nov. |
Trimma meranyx View in CoL n. sp.
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Suggested common name: Day-night pygmy goby.
Trimma View in CoL species 3 – Allen & Erdmann, 2012: 946 (West Papua, Indonesia and Vanuatu).
Material Examined. Five lots, 9 specimens ( 14.8–18.7mm SL, all females) plus one specimen for genetic analysis, from Lembeh Strait, Sulawesi ( type locality), Raja Ampat, the Fakfak Peninsula, and the Milne Bay district, Papua New Guinea.
Holotype. ROM 94170, 18.7 mm SL female, Indonesia, Sulawesi, Lembeh Strait, Nudi Retreat, ( 01° 29.190' N, 125° 14.464'E), 45–55 m, clove oil, 0 6 Jan., 2013, M.V. Erdmann.
Paratypes. MZB.22092, 2 (14.8–15.2), collected with the holotype. ROM 94721, 1 (17.8), collected with the holotype. ROM 94017, 1 (18.0), Indonesia, Sulawesi, Lembeh Strait, Nudi Retreat ( 01° 21.190’ N; 124° 37.757' E), 65 m, 8 Sept., 2012, M. V. Erdmann. ROM T12753, 1 (11.7), collected with ROM 94017, tissue specimen.
Non-type Material. ROM 94174, 2(18.4–18.5), Indonesia, Raja Ampat, Fam Is, Twin Towers, ( 00° 35.454' S, 130° 18.012' E), 70 m, clove oil, 0 4 Feb., 2013, M. V. Erdmann. ROM 94502, 1 (17.9), PNG, Milne Bay District, Wahoo Pt, ( 10° 15.177' S, 150° 47.048' E), 70 m, clove oil, 14 June, 2013, M.V. Erdmann. WAM P.33090.005, 1 (16.9), Indonesia, West Papua, N side of island directly S of the city of Fakfak, ( 02° 58.525’ S, 132° 16.715’ E), 58– 62 m. G.R. Allen & M.V. Erdmann, 21 Mar., 2009.
Diagnosis. Trimma meranyx has a narrow bony interorbital (<55% pupil width), 8–9 scales in the predorsal midline, usually cycloid scales along the upper border of the opercle which may be in up to three horizontal rows, two (usually cycloid) cheek scales, a very slightly elongate second dorsal spine usually reaching posteriorly to between the base of the spine and the base of the second ray of the second dorsal fin, rays of the pectoral fin unbranched, a fifth pelvic fin ray which branches once dichotomously and is 64–85% the length of the fourth ray, and a full basal membrane connecting the inner margins of the fifth pelvic fins rays in undamaged specimens. The black posterior half of the caudal peduncle with a large white spot dorsally and ventrally just anterior to the procurrent caudal fin rays is unique within the genus.
Description. Dorsal fins VI + I 8, second spine only slightly elongate, reaching posteriorly to between the bases of the spine and ray 2 second dorsal fin when adpressed, rays all branched except posterior element of last ray, last ray reaching posteriorly 22 –38% of distance from its base to first dorsal procurrent ray; anal fin I 8, first ray branched or unbranched, posterior element of posteriormost ray unbranched; posteriormost ray 24 –40% of distance from its base to first ventral procurrent ray; pectoral fin 15 (once 14), all rays unbranched, fin reaching posteriorly to a vertical line between anus and anal-fin origin; pelvic fin I 5, no frenum, basal membrane 100 % length of inner branch of fifth ray when undamaged, first four rays with one sequential branch, fifth ray branched once dichotomously and 74– 83 –85% length of fourth, fourth ray reaching posteriorly to between urogenital papilla and base of anal ray 2. Lateral scales usually 23, when 24, a small extra scale present in axil of pectoral fin, and usually only on one side of body (n = 2, including left side of holotype); anterior transverse scales 7– 8 –9 (mean = 8); posterior transverse scales 7 (once 6); predorsal scales 8– 9 (mean = 8.4); two usually cycloid scales on posterodorsal part of cheek (ctenoid in 2) lying above sensory papillae line b; opercle with 2– 3 –4 horizontal rows of 3, 2 and 1 usually cycloid scales, scales in upper row or rows may be ctenoid in larger specimens (> 17 mm SL); pectoral fin base with 2– 3 vertical rows of cycloid scales with 3 scales in posterior row; 6– 7 cycloid prepelvic scales (in midline anterior to basal membrane); 12 circumpeduncular scales; body scales ctenoid except for cycloid scales on anterior belly midline, and beneath pectoral fin base; body scales extend anteriorly to just behind eye. Gill opening extending anteroventrally to a vertical below mid-pupil. Teeth not examined in detail, but appear to be typical of other Trimma . Cephalic sensory papillae ( Fig. 1 View FIGURE 1 ) row counts given in Table 1 View TABLE 1 . Tongue truncate with broadly rounded edges. Gill rakers on first arch 3 –4 + 13– 14 = 16– 17. Anterior nasal opening a short tube extending out over upper lip, posterior nasal opening a pore with a raised rim, both protruding from slightly raised oval sac confined to anterior half of snout. Bony interorbital 40– 43 –54% pupil width, with a slight furrow between the interorbital papillae and no postorbital groove. No ridge of tissue (or dermal crest) extending anterior to first dorsal spine. Epaxialis extending anteriorly to above a vertical with posterior margin of pupil. Vertebral transition not examined, but predicted to be Type B (see Winterbottom, 2003, Fig. 14, inset, and Winterbottom & Zur, 2007, Fig. 3 View FIGURE 3 B)
Colour pattern (live, from three images of specimens from Indonesia, Papua New Guinea, and the Philippines). The specimen from Lembeh Strait, Sulawesi (ROM T12753, 11.7 mm SL; Fig. 2 View FIGURE 2 A) has a light greenish-yellow head, this colour becoming gradually darker and greener posteriorly on the body, and there are numerous randomly scattered small dark spots on the body. About half-way along the peduncle, the colour becomes dark purple-red, grading posteriorly to reddish black before lightening to a red flush over the base of the caudal fin rays. An almost pupil-diameter sized white saddle lies across the dorsal surface of the peduncle with its posterior edge just anterior to the procurrent fin rays, and a similar spot is present across the ventral midline. The spots are separated by a onescale width band of reddish black in the lateral midline. A similar, but somewhat smaller, pair of white saddles is present at about the point where the sides of the peduncle start to darken. There is a basal, half-pupil width dark band in both dorsal and anal fins, containing numerous scattered iridocytes. This is followed by a broad dirty yellow band in the middle of the fins, with another diffuse dark stripe distally (again with iridocytes in the anal but not the dorsal fin). All visible fin rays are red, and the iris is plain yellow. Live specimens from Mactan and Milne Bay are essentially similar in the details that are visible, although the body is more yellow with only a slight greenish cast. A freshly collected 18.5 female (ROM 94174; Fig. 2 View FIGURE 2 B) from Fam Is, Raja Ampat is more brightly coloured with a yellow abdominal region, the posterior half of the peduncle dark red preceded by a diffuse lighter band, there is no trace of the anterior pair of white saddles, and the distal tip of the anterior part of the first dorsal fin is white. The caudal fin (posterior to the basal red flush) is yellow margined above and below by diffuse white/ blue stripes. The rays and membranes of the pelvic fin are yellow, the extreme tips of the third and fourth rays are tipped with white. A freshly collected Vanuatu specimen is yellow with little or no trace of green over the head and body. The posterior half of the peduncle is diffuse black grading to reddish on the caudal fin base, and there is no trace of an anterior pair of white saddles. A live specimen photographed at the type locality ( Fig. 2 View FIGURE 2 C) has a greenish-yellow head, and the scale margins on the body appear to be faintly outlined with dark red.
Colour pattern in alcohol. Background colour pale straw yellow. Head, body, and upper half of pectoral fin base with a liberal scattering of melanophores and brown chromatophores, which also faintly outline the scale margins on the upper body. Pigmentation of the posterior half of the caudal peduncle intensified, with larger brown chromatophores concentrated at the dorsal and ventral margins, and smaller brown chromatophores more concentrated on the mid-lateral side of the peduncle ( Fig. 3 View FIGURE 3 ). This pigmentation continues posteriorly onto the bases of the caudal fin rays. It is interrupted by a pair of eye-diameter width, unpigmented saddles dorsally and ventrally, with the posterior margins of the saddles reaching to the anterior procurrent caudal fin rays. The midlateral septum from below the origin of the second dorsal fin with a dotted line of elongate melanophores. A half-pupil width band of very dark brown chromatophores is present at the bases of the two dorsal fins, which expands in width posteriorly to fill the fin membrane between the last two dorsal rays. A similar basal band is present in the anal fin, but there is also a distal band of melanophores which grade into dark brown melanophores posteriorly. The caudal fin has a few scattered melanophores and a pupil-diameter width diffuse distal stripe made up of melanophores. The membranes of pectoral and pelvic fins are essentially hyaline (there may be a few scattered pigment cells). Anterior half of lips with scattered melanophores and brown chromatophores. There is some variation in the intensity of the pigmentation of the posterior half of the caudal peduncle between specimens (cf Fig. 3 View FIGURE 3 A and 3B), and in several specimens almost all the pigmentation appears to consist of chromatophores, with only a few melanophores.
Comparisons. Trimma meranyx belongs to a limited grade of three described species which share a narrow bony interorbital (<80% pupil width), lack an elongate spine in the first dorsal fin, have a scaled predorsal midline, cheek scales and a scaled opercle, and only unbranched rays in the pectoral fin rays. The described species are: Trimma emeryi Winterbottom, 1985 , T. fasciatum Suzuki, Sakaue & Senou, 2012 , and T. flavatrum Hagiwara & Winterbottom, 2007 . The first two of these species lack the dark posterior half of the caudal peduncle, but such a feature is present in T. flavatrum , which differs in having a shorter fifth pelvic fin ray (40–60% fourth, vs. 74–85%), lacking a basal membrane (vs. full membrane present), and in lacking any indication of white saddles on the caudal peduncle (vs. present). A pair of white saddles over the posterior peduncle is present in the western Indian Ocean T. bisella Winterbottom, 2000 , but that species lacks scales in the predorsal midline (vs. present), has 9–10 dorsal rays (vs. 8), has an elongate 2nd dorsal spine (vs. not elongate), an unbranched fifth pelvic ray with no basal membrane (vs. branched and full respectively), and lacks scales on the cheek and opercle (vs. both present).
Distribution. Trimma meranyx is currently known from specimens from Lembeh Strait, Raja Ampat, the Fakfak Peninsula, and the Milne Bay district, Papua New Guinea at depths of 45– 70 m. There are also photographs of what appears to be this species from Mactan, Philippines and from Vanuatu.
Etymology. The name is a compound noun derived from the Greek words “mera”, meaning day, and “nyx”, meaning night, and thus translates to the ‘day-night pygmy goby’ in reference to the unique colouration of a black peduncle with twin “suns” dorsally and ventrally. The name is to be treated as a noun in apposition. This species has been informally referred to as Trimma RW sp. 91.
Discussion. Given recent studies using the Cytochrome Oxidase 1 gene (Winterbottom et al., 2014) that suggest that morphology may only be distinguishing about half the genetic species of this genus, and that more than one exceedingly similar morphological species can be found in the same area (and even in the same collection), we prefer to err on the side of caution in selecting type specimens, and accordingly have restricted the types to specimens from the Lembeh Strait, Sulawesi. We note that the anterior pair of white saddles are not evident in the preserved material available to us. Although our own photographs of freshly collected specimens do show evidence of these saddles, they are not as obvious as in live specimens, and we see no trace of them in the photograph of the specimen from Vanuatu. It is not known whether the Vanuatu specimen represents the same species as we describe here.
Trimma meranyx is a "twilight zone" species, with most specimens reported from the 70 m depth range (though they have been collected as shallow as 45–55 m in the Lembeh Strait). They do not appear to school or aggregate as with many congeners, but rather are found individually in small caves and crevices, frequently quite deep into the interstices of the reef and only visible with concerted effort using a strong underwater flashlight. All specimens collected by the second author were found exclusively on deep reefs sheltered from wave exposure but nonetheless exposed to currents, and although there was frequently significant silt build-up present in the caves in which they sheltered, water visibility was always good as a result of significant water flow over the reefs.
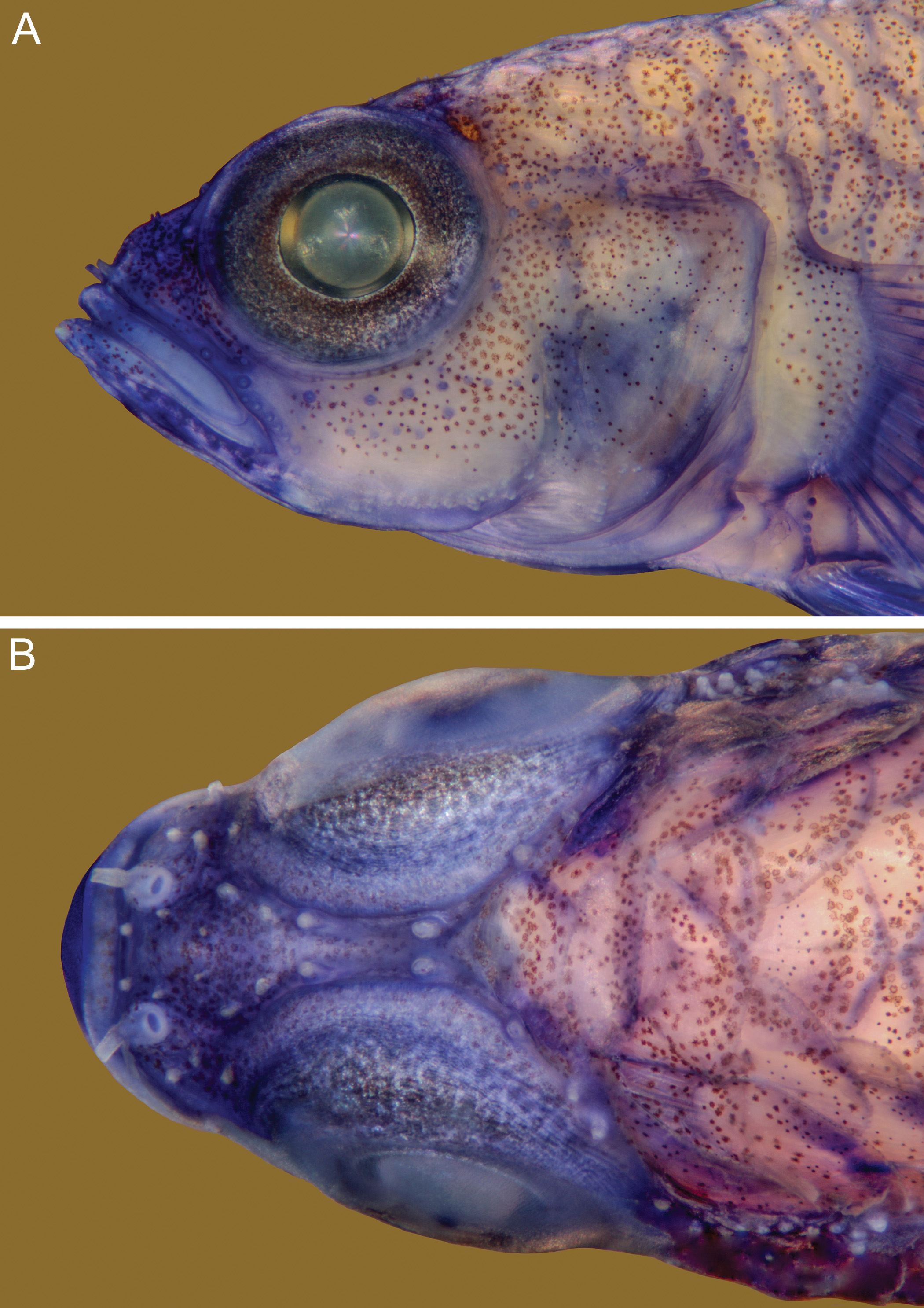


TABLE 1. Counts of cephalic sensory papillae in the different rows of the three new species of Trimma. Data given as minimum to maximum values, with the mean, Standard Deviation and number of specimens examined respectively given in parentheses, except where the value was invariant, where only the number of specimens is given. Terminology follows that used in Winterbottom (2011).
| Papillae row | T. meranyx | T. pajama | T. zurae |
|---|---|---|---|
| Row a | 5 (9) | 5 (11) | 5 (6) |
| Row b | 3–5 (4, 0.5, 9) | 3–4 (3.1, 0.29, 11) | 6–8 (7, 0.82, 6) |
| Row c | 6 (9) | 5 (11) | 5–6 (5.8, 0.37, 6) |
| Row cp | 1 (9) | 1 (11) | 1 (6) |
| Row d | 4–6 (4.9, 0.57, 9) | 2–4 (3.1, 0.51, 11) | 7–9 (7.5, 0.76, 6) |
| Row d' | 6–7 (6.4, 0.47, 9) | 3–5 (3.5, 0.66, 11) | 6–9 (7.2, 0.9, 6) |
| Row e –ant. | 10–13 (11.6, 0.96, 9) | 7–10 (8.4, 0.8, 10) | 11–16 (13, 1.83, 6) |
| Row e –post. | 10–14 (12.6, 1.34, 9) | 8–11 (9.4, 0.92, 10) | 12–17 (14.2, 1.57, 6) |
| Row i – ant. | 6–7 (6.8, 0.42, 9) | 6–7 (6.3, 0.45, 11) | 7–8 (7.5, 0.5, 6) |
| Row i – post. | 6–8 (7.0, 0.47, 9) | 6–7 (6.5, 0.50, 11) | 7–8 (7.2, 0.37, 6) |
| Row p | 8 (9) | 6–7 (6.8, 0.39, 11) | 6 (6) |
| Row r | 1–2 (1.1, 0.31, 9) | 1–2 (1.9, 0.29, 11) | 2 (6) |
| Row f | 3–4 (3.1, 0.21, 9) | 2–3 (2.6, 0.50, 9) | 3–4 (3.2, 0.37, 6) |
| Row cs" | 3 (8) | 3 (11) | 3 (6) |
| Row g | Apparently absent | 2 (1) | 6–10 (7.2, 1.47, 5) |
| Row x | 7–9 (7.4, 0.68, 9) | 6–8 (7.0, 0.63, 5) | 7–9 (7.7, 0.75, 6) |
| Row z | 5–9 (6.5, 1.38, 6) | 4–6 (5.4, 0.80, 5) | 6–7 (6.3, 0.47, 6) |
| Row ot | 8–11 (9.8, 1.09, 8) | 8–11 (9.2, 1.07, 6) | 12–15 (12.8, 1.07, 6) |
| Row os | 5–7 (6.3, 0.83, 4) | 3–4 (3.7, 0.47, 3) | 5–7 (5.6, 0.8, 5) |
| Row oi | 3–4 (3.8, 0.43, 8) | 2–3 (2.7, 0.45, 7) | 4–5 (4.2, 0.37, 6) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Trimma meranyx
| Winterbottom, Richard, Erdmann, Mark V. & Dita Cahyani, N. K. 2014 |
Trimma
| Allen 2012: 946 |