
Oribatella hungarica Balogh, 1943
|
publication ID |
https://doi.org/10.11646/zootaxa.4857.1.8 |
|
publication LSID |
lsid:zoobank.org:pub:D381069A-B251-4896-91AF-C23C37D81BE5 |
|
DOI |
https://doi.org/10.5281/zenodo.4396593 |
|
persistent identifier |
https://treatment.plazi.org/id/03E087E9-FFFC-FFC7-FF66-FCC9C5DEDBB7 |
|
treatment provided by |
Plazi |
|
scientific name |
Oribatella hungarica Balogh, 1943 |
| status |
|
Oribatella hungarica Balogh, 1943 View in CoL
Morphology of adult
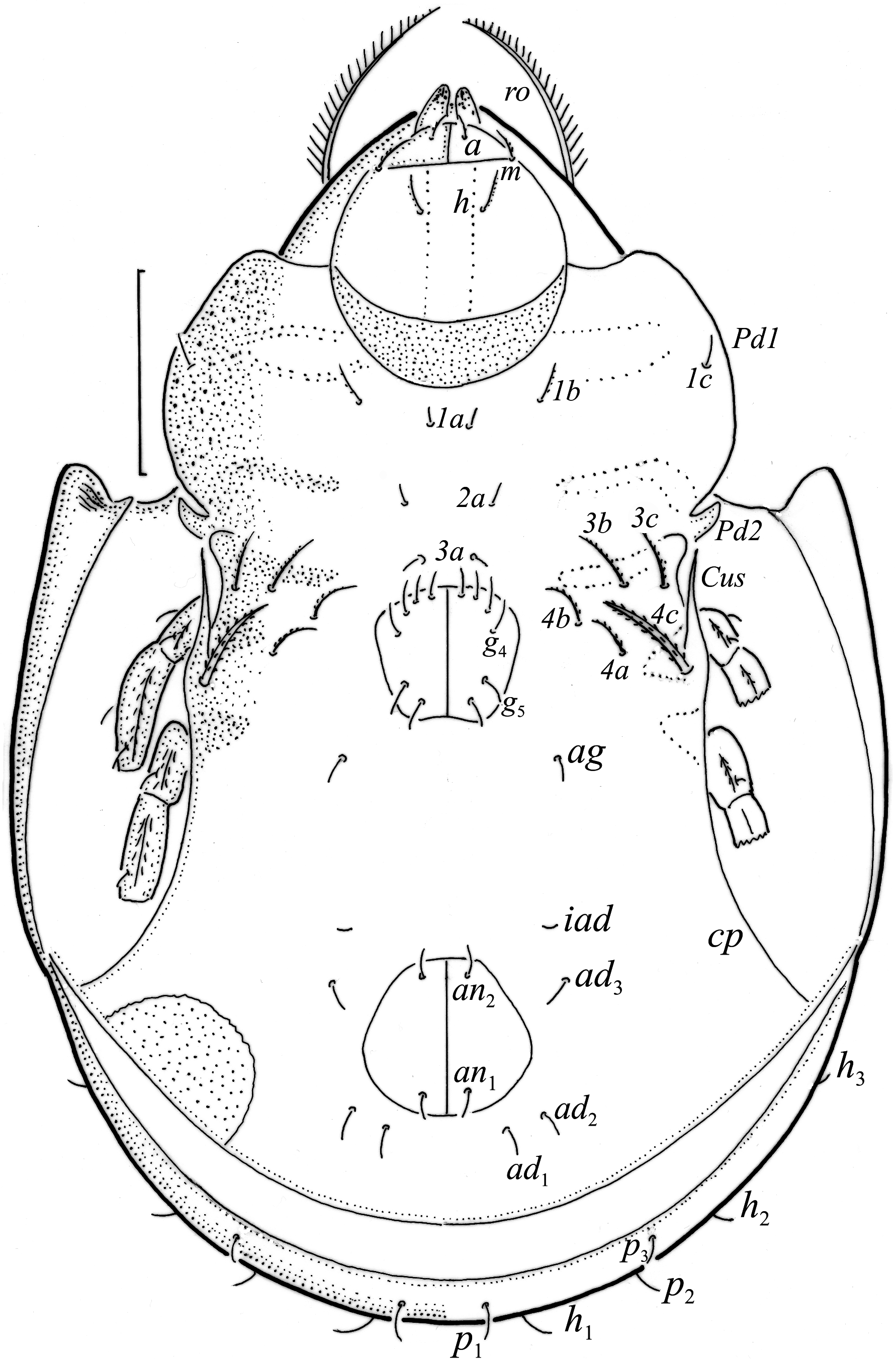
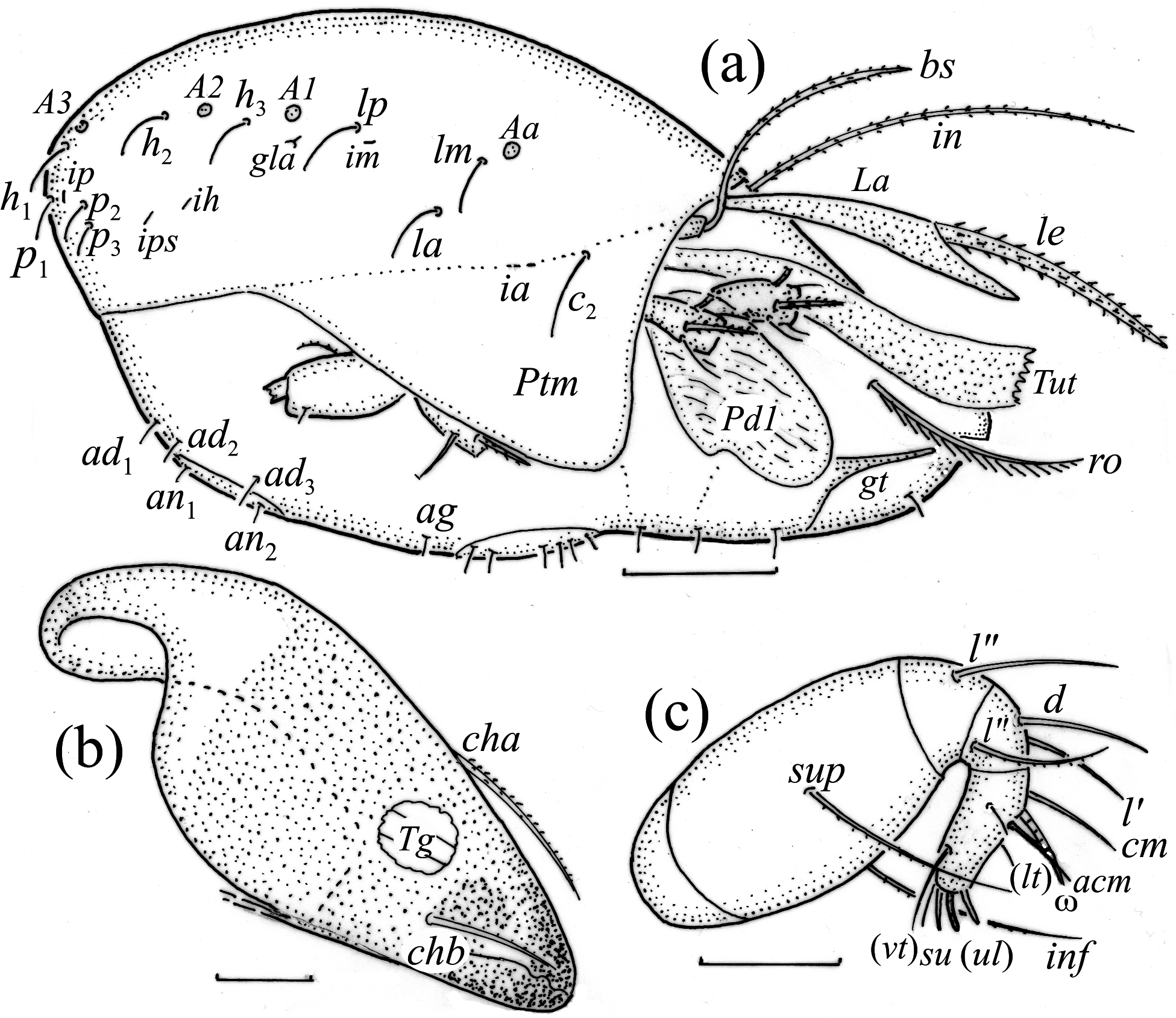
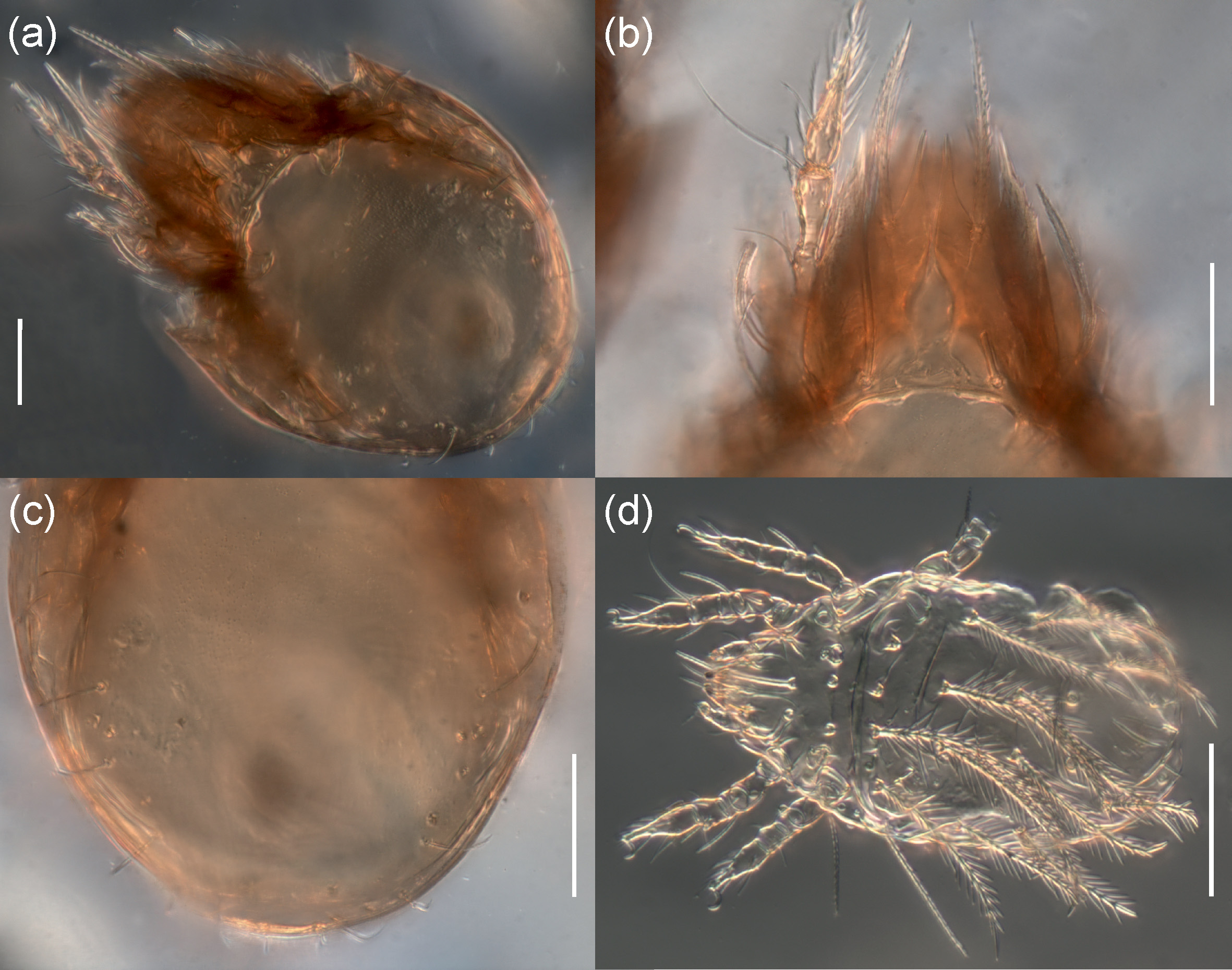
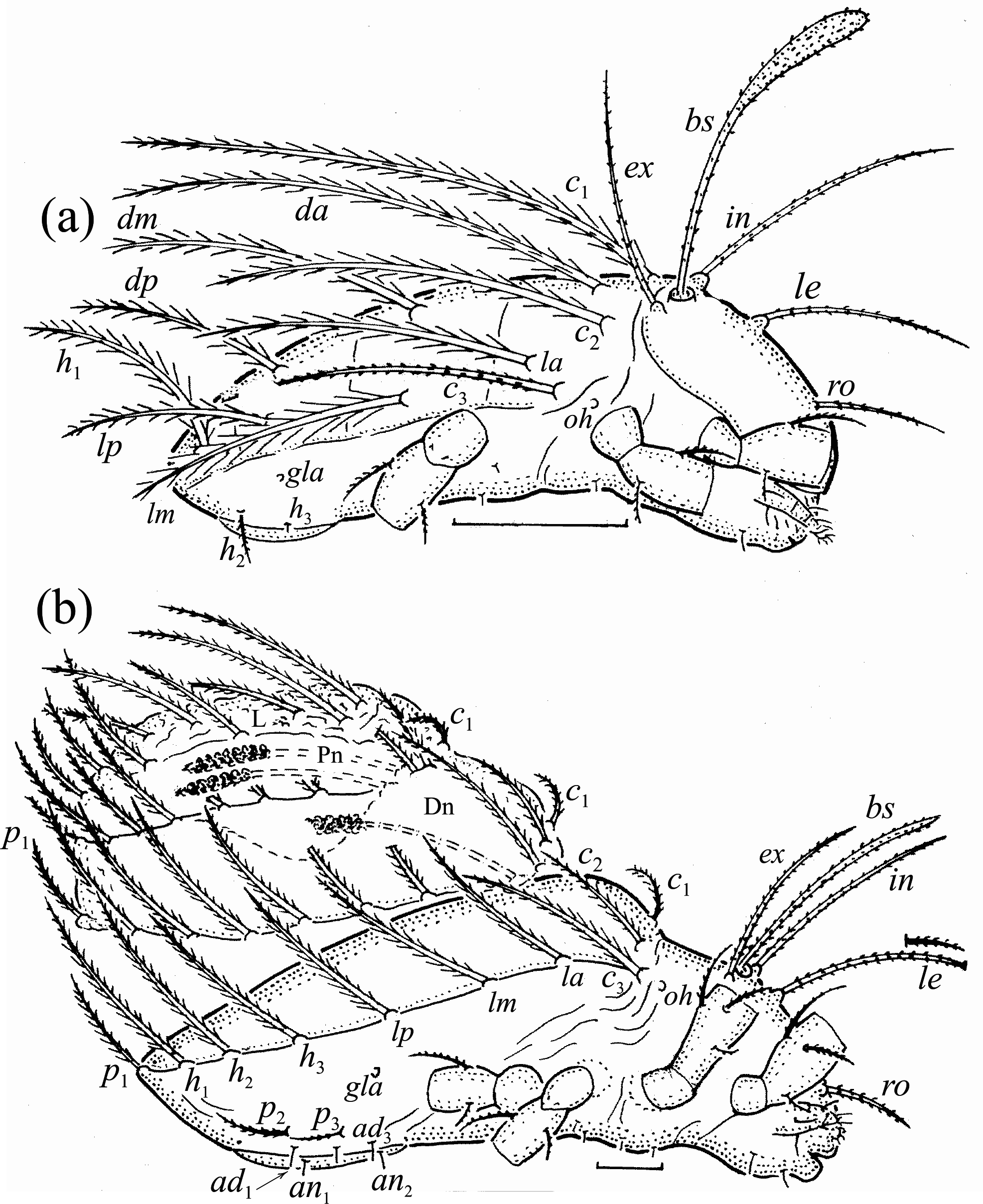
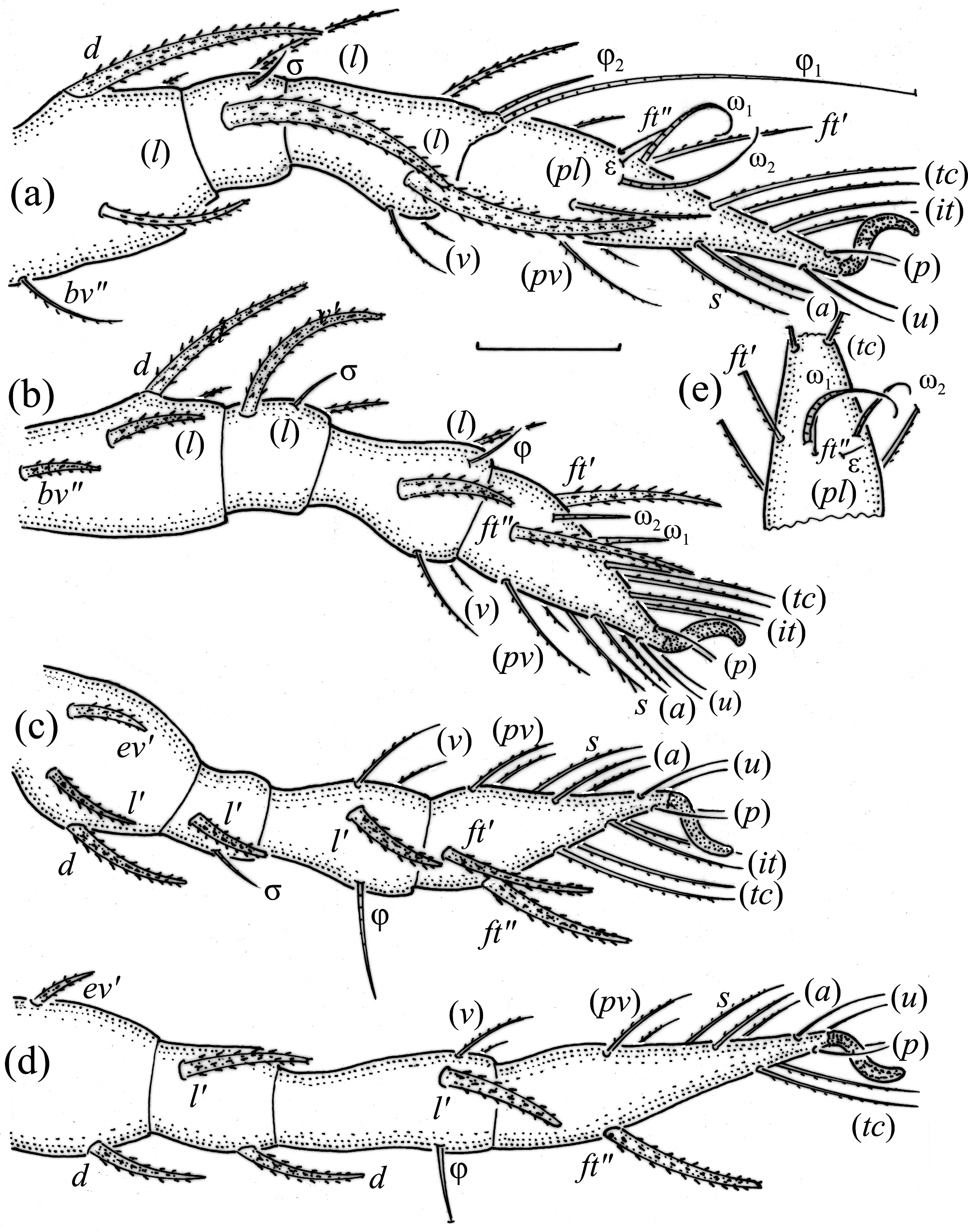
Adults studied here ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3a View FIGURE 3 , 4 View FIGURE 4 a–c, 5) similar to those investigated by Balogh (1943) and Bernini (1978), but see Remark below. Mean length (and range) of females 311.3±8.87 (295–337, N=100) and males 294.4±6.38 (range 277–307, N=50); mean width of females 201.5±5.94 (187–217) and males 189.2±6.11 (175–205). Subcapitular se- tae h and m longer than a, all barbed ( Fig. 2 View FIGURE 2 ). Chelicera chelate, seta cha longer than chb, both barbed ( Fig. 3b View FIGURE 3 ). Palp relatively short, palpal setae sup and inf longer than other setae, sup, inf and l” on tibia barbed, other setae smooth ( Fig. 3c View FIGURE 3 ). Formula of palp setae [trochanter to tarsus (+ solenidion ω)]: 0-2-1-3-9(1). Epimeral setae 1a, 1c, 2a and 3a short ( Fig. 2 View FIGURE 2 ) and smooth, seta 1b slightly longer, setae 3b, 3c, 4a and 4b clearly longer and barbed, and seta 4c longest and thickened; all barbed. Most leg setae barbed, seta l” on genu I and II and tibia I and II, and l’ on genu III and genu and tibia IV thickened ( Fig. 5 View FIGURE 5 , Table 2). Formulae of leg setae [trochanter to tarsus (+ solenidia), Table 3)]: I—1-5-3(1)-4(2)-20(2); II—1-5-3(1)-4(1)-15(2); III—2-3-1(1)-3(1)-15; IV—1-2-2-3(1)-12. Legs bidactylous.
Remark. The adults of O. hungarica investigated here are smaller than those investigated by Bernini (1978; length of females 330–390, males 290–390; width of females 255–260, males 195–255), but other characters are similar as in Balogh (1943) and Bernini (1978).
Diagnosis of juveniles
Juveniles light-brown and nymphs apopheredermous, i.e. they carry exuvial scalps of previous instars away from dorsal integument, using modified setal pair da. Larva with 12 pairs of gastronotal setae, most with long barbs, except for c 3 with short barbs and minute h 3, nymphs with 13 pairs of these setae, with short barbs ( dm and dp are lacking). In tritonymph, leg seta d on all femora and genu IV, l” on femora I and II and pair l on genua and tibiae I and II, l’ on femur III and genua and tibiae III and IV, and ft on tarsi II–IV thicker than other setae on these segments. In all juveniles, humeral organ present.
Description of juvenile stages
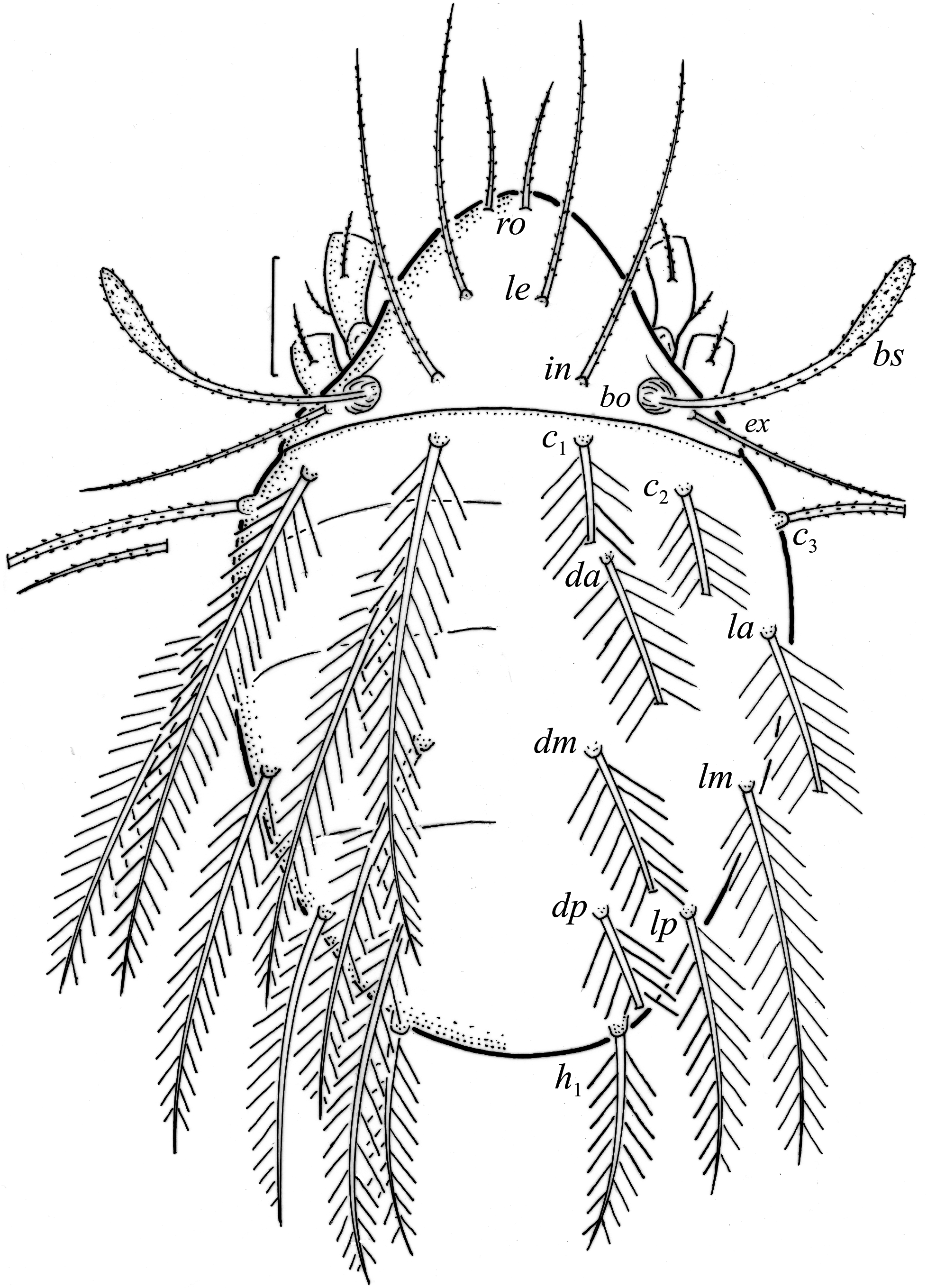
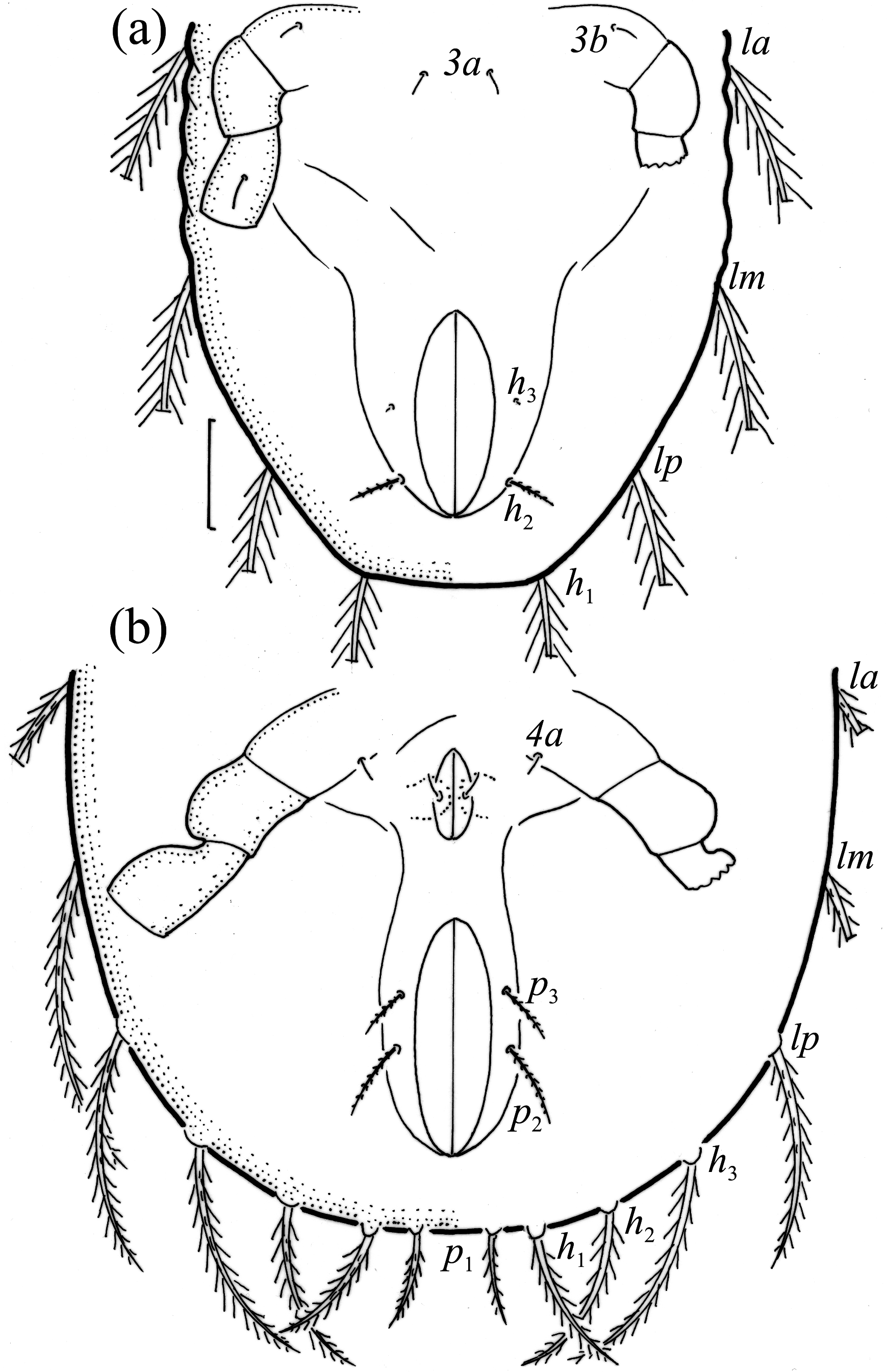
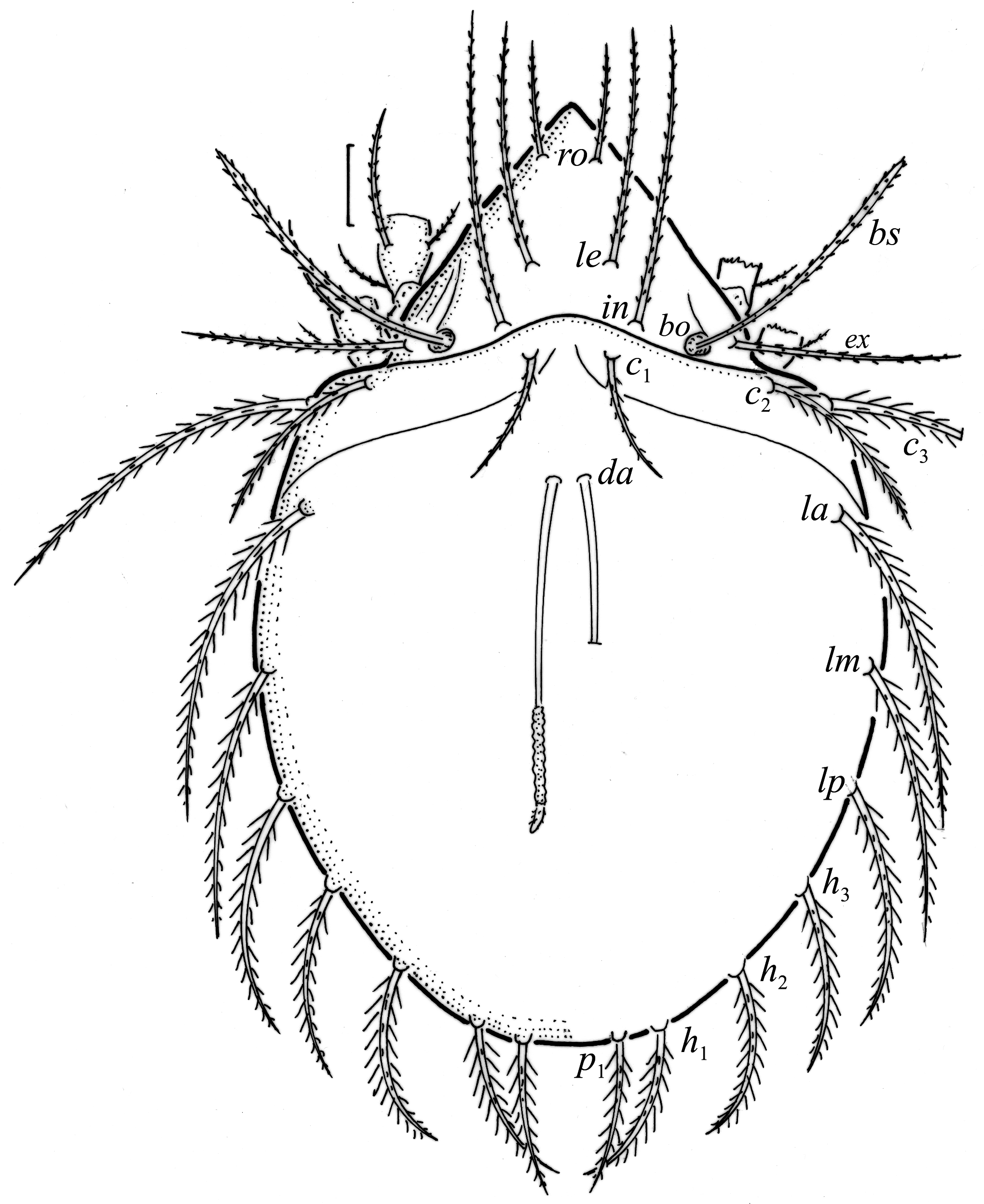
Larva oval in dorsal aspect ( Figs. 4d View FIGURE 4 , 6 View FIGURE 6 ) and light brown. Prodorsum subtriangular, all prodorsal setae long, with short barbs, in longest, ro shortest ( Table 4). Mutual distance between setal pairs le about two times longer than between pair ro, and between pair in about three times longer than between pair ro. Seta le inserted midway between pairs ro and in. Bothridium rounded, bothridial seta narrowly clavate and barbed.
Gastronotum of larva with 12 pairs of setae, including minute h 3 inserted laterally to medial part of anal valves ( Figs. 6 View FIGURE 6 , 7a View FIGURE 7 , 8a View FIGURE 8 ). Most gastronotal setae long, with long barbs, except for long c 3 and medium sized h 2 with short barbs, and minute h 3; length of setae decreasing from anterior to posterior ( Figs. 6 View FIGURE 6 , 8a View FIGURE 8 , Table 4). Anal valves (seg- ment P) glabrous. Cupules ia, im, ip and ih not observed, gland opening lateroventral to seta lp. Humeral organ anterior to seta c 3 present ( Fig. 8a View FIGURE 8 ). Most leg setae barbed, l” on genu and tibia I and genu II, and l’ on genu III longer than other leg setae ( Fig. 9 View FIGURE 9 ).
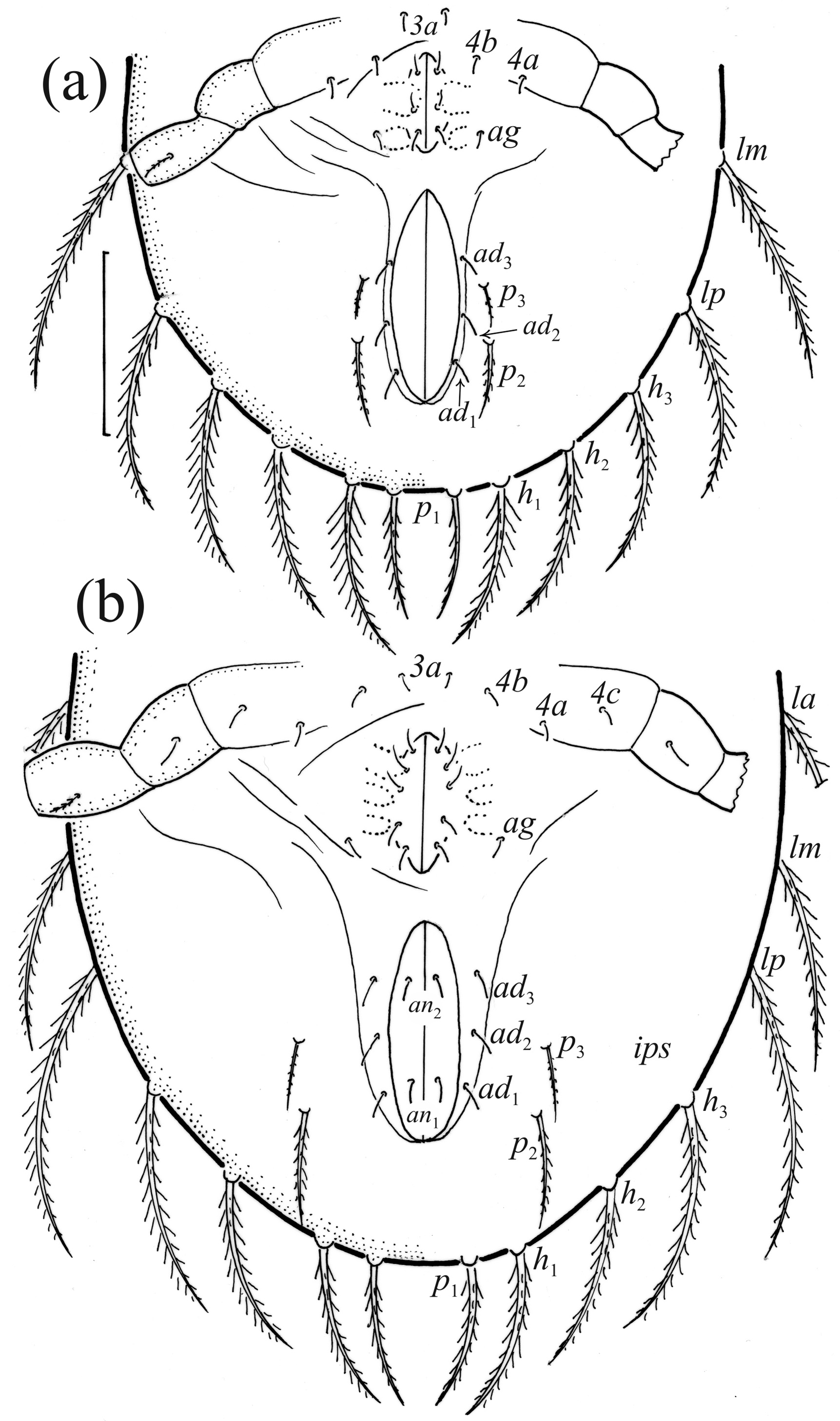
Nymphs more stocky than larva, with setiform bothridial seta, but in protonymph slightly clavate, length and distribution of prodorsal setae as in larva. Gastronotum of protonymph with 13 pairs of setae because setae of p - series appearing ( Fig. 7b View FIGURE 7 ), and remaining in deutonymph and tritonymph ( Figs. 10a, 10b View FIGURE 10 ), but setae of dm and dp lost and remaining absent in other nymphs. In all nymphs, most gastronotal setae long ( Table 4), except for medium sized p 2 and p 3; p 2 slightly longer than p 3. Setal pair da long, smooth, inserted close to each other and posterior to setal pair c 1, apical end of da adapted for bearing exuvial scalps of previous instars. Other gastronotal setae in marginal position on gastronotum and barbed, but setae p 2 and p 3 with shorter barbs than other setae. All nymphs carry exuvial scalps of previous instars ( Fig. 8b View FIGURE 8 ), using long setal pair da. In protonymph, one pair of setae appearing on genital valves, and two pairs added in deutonymph and tritonymph each ( Figs. 7b View FIGURE 7 , 10a, 10b View FIGURE 10 ); all short and smooth. In deutonymph, one pair of aggenital setae and three pairs of adanal setae appearing, and present in tritonymph; all short and smooth. Anal valves of protonymph (segment AD) and deutonymph (segment AN) glabrous, but in tritonymph two pairs of anal setae present, all short and smooth. Cupules ia, im, ip, ih, ips and iad not observed, opisthonotal gland opening as in larva ( Figs. 7b View FIGURE 7 , 8b View FIGURE 8 , 10a, 10b View FIGURE 10 , 11 View FIGURE 11 ). In tritonymph, leg setae d on all femora and genu IV, l” on femora I and II and pair l on genua and tibiae I and II, l’ on femur III and genua and tibiae III and IV, and ft on tarsi II–IV thicker than other setae on these segments ( Fig. 12 View FIGURE 12 , Table 2). Solenidia ω 1 and ω 2 on tibia I curved outwards. In all nymphs, humeral organ as in larva.
Summary of ontogenetic transformations
In all instars of O. hungarica , the prodorsal setae are long, and seta in is longest and ro is shortest, but in the juveniles le is clearly longer than ro, whereas in the adult these setae are of similar length, and le is clearly thicker than other setae. In all juveniles, seta ex is long, whereas in the adult it is short. The opening of bothridium is rounded in all instars, but in the adult it gains small scales. In the larva, the bothridial seta is narrowly clavate, whereas in other instars it is fusiform. The larva has 12 pairs of gastronotal setae, including minute h 3, the nymphs have 13 pairs ( p - series appears, dm, dp are lost), whereas the notogaster of adult loses setae c 1, c 3 and da, such that 10 pairs of setae remain. The formula of gastronotal setae is 12-13-13-13-10 (from larva to adult), and formulae of epimeral setae are 3-1-2 (larva, including scaliform 1c), 3-1-3-1 (protonymph), 3-1-3-2 (deutonymph) and 3-1-3-3 (tritonymph and adult). Formula of genital setae is 1-3-5-6 (protonymph to adult), aggenital setae is 1-1-1 (deutonymph to adult), and formula of segments PS−AN is 03333-0333-022. The ontogeny of leg setae is shown in Table 3.
Distribution, ecology and biology
Oribatella hungarica View in CoL has a Mediterranean distribution (Subías 2020) and is considered sylvicolous ( Mahunka & Mahunka-Papp 2004, Ivan & Vasiliu 2006). This species was described from Hungary ( Balogh 1943), and recorded from Italy ( Bernini et al. 1995), southern Austria ( Krisper et al. 2017), Hungary ( Mahunka & Mahunka-Papp 2004), Bulgaria ( Csiszár & Jeleva 1962) and Romania ( Ivan &Vasiliu 2000). Oribatella hungarica View in CoL is frequent and rela- tively abundant in litter and moss of mesophilous forests in hilly regions ( Ivan & Vasiliu 2000). This species tolerated the cement dust pollution in beech forests in Romanian Southern Subcarpathians ( Ivan & Vasiliu 2012), and also chemicals used against defoliating insects in oak forests (Ivan 2009), but is sensitive to the forest soil pollution with heavy metals and fluorine ( Ivan & Vasiliu 2009).
In this study, O. hungarica View in CoL was present in the upper soil horizon of different oak forests ( Table 1), with the highest density in Roşcani (26.8 individuals per 500 cm 3), and lowest in Heltiu (7.8 individuals per 500 cm 3). In all soils, the adults highly dominated the juveniles, which constituted 1–11% of sample populations or were absent. In the soil in Ampoiţa, all instars of O. hungarica View in CoL were present, and the stage structure of this species was the following: one larva, one protonymph, two deutonymphs, three tritonymphs and 41 adults.
In these soils, females of O. hungarica were more abundant than males, the sex ratio (females: males) varied (1:0.43–1:0.58), and 15–58% of females were gravid ( Table 5). The gravid females carried 1–2 relatively large eggs, each 163–169 x 102–108, which constituted about 53% of the total body length of females.
In most oak forests, the differences of body length and width of females and males of O. hungarica were indistinct. Only in Heltiu, females were significantly longer than in Ampoiţa and Roşcani, and only in Roşcani males were significantly shorter than in other forests. In Ampoiţa, females were significantly wider than in Copălău and Heltiu, and in Urecheşti males were significantly wider than in Roşcani, Copălău and Ampoiţa ( Table 5). In all for- ests, females of O. hungarica were significantly longer and wider than males.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Oribatida |
|
Family |
|
|
Genus |
Oribatella hungarica Balogh, 1943
| Seniczak, Stanisław, Ivan, Otilia, Marquardt, Tomasz & Seniczak, Anna 2020 |
Oribatella hungarica
| Balogh 1943 |
Oribatella hungarica
| Balogh 1943 |
Oribatella hungarica
| Balogh 1943 |
Oribatella hungarica
| Balogh 1943 |
Oribatella hungarica
| Balogh 1943 |
Oribatella hungarica
| Balogh 1943 |
Oribatella hungarica
| Balogh 1943 |
Oribatella hungarica
| Balogh 1943 |
Oribatella hungarica
| Balogh 1943 |
O . hungarica
| Balogh 1943 |
O . hungarica
| Balogh 1943 |