
PHLOEIDAE Amyot & Serville, 1843
|
publication ID |
https://doi.org/10.1111/j.1096-0031.2008.00224.x |
|
DOI |
https://doi.org/10.5281/zenodo.4334450 |
|
persistent identifier |
https://treatment.plazi.org/id/03E187AB-6B75-FFF0-FCFD-FA5E104E4937 |
|
treatment provided by |
Valdenar (2020-07-31 15:01:18, last updated 2024-11-26 07:01:54) |
|
scientific name |
PHLOEIDAE Amyot & Serville |
| status |
|
PHLOEIDAE Amyot & Serville (+ SERBANINAE Leston), sensu Distant
Historical: Amyot and Serville (1843) first recognized the neotropical ‘‘Phleides’’ as a suprageneric taxon. Stål
(1872) treated the group as a subfamily of Pentatomidae . Leston (1953b) and Lent and Jurberg (1965) monographed the two genera and three species. Rolston and McDonald (1979) treated the group as a family. In Gapud̕s (1991) analysis, the Phloeidae came out in a basal position following the Urostylididae . The interpretation of at least seven characters in Gapud̕s matrix should be changed, based on more thorough morphological observations. Apparently, Gapud (1991) exam-
ined specimens of Phloea corticata (Drury) , and made the same erroneous interpretations of the female genitalia found in Lent and Jurberg (1965) and Rolston and McDonald (1979). Thegonocoxites 9 (= 2nd gonocoxae,
2nd valvifers) of these authorsare infactthe laterotergites 9, whereas the 9th paratergites ( Rolston and McDonald, 1979) correspond to the laterotergites 8 (= 8th paratergites). Character 13 (2nd valvifers) of Gapud (1991), coded as distinctly separate, should be changed to 2nd valvifers with a distinct median fusion line. Also, the Phloeidae (including Serbaninae) have a membranous gonangulum rather than this structure being well sclerotized, as suggested by Gapud (character 17).
The Serbaninae, originally described in the Phloeidae by Distant (1906), is monotypic and restricted to Borneo. Serbana borneensis Distant was removed and established as a higher taxon within the Pentatomidae by Leston (1953b), based mostly in the morphology of the male genitalia; this placement has more recently been accepted by Rider (2006).
Analytical result: Phloeidae sensu lato receives support from our morphological analyses, on the basis of the invariant characters: body foliations present (31) and eyes divided on sagittal plane (81); and the homoplasious characters: base of corium expanded (201) and spiracles of segment VIII present, concealed by segment VII (391). This is despite the fact some prior authors have rejected the association of Phloeidae sensu stricto with Serbana . Nonetheless, the novel morphology of the Phloeidae complicates its placement within the Pentatomoidea on the basis of morphology alone. In the total evidence analysis of 92 taxa under an equal-cost regime, Phloeidae (including Serbana ) becomes the sister group of a clade including the Plataspididae + Cyrtocoris + Megaris . The inclusion of Crytocoris in this grouping is found nowhere else in our analyses, and we consider it to be an artifact relating to the large amount of missing data in the 92-taxon combined analysis. Cyrtocoris is ‘‘correctly’’ placed as the sister group of the Pentatomidae under 1: 2, 2: 1, and 2: 2 cost regimes. At the same time, the Phloeidae move into the clade containing the Dinidoridae , Tessaratomidae , and some of the ‘‘cydnoid’’ taxa ( Figs 54 View Fig and 55 View Fig ). That result is similartothecombinedmolecularanalysesundera 2: 2 weighting scheme ( Fig. 46 View Figs 45–48. 45 ) and the 52-taxon total evidence analysis under a 1: 1 weighting scheme ( Fig. 51 View Figs 49–52. 49 ). Further sequencing may or may not help to resolve this issue.
Amyot, C. J. B., Serville, A., 1843. Histoire naturelle des insectes He mipte`res. Librairie Encyclope'dique de Roret, Paris.
Distant, W. L., 1906. Oriental Heteroptera. Ann. Soc. Entomol. Belg. 50, 405 - 408.
Gapud, V., 1991. A generic revision of the subfamily Asopinae with consideration of its phylogenetic position in the family Pentatomidae and superfamily Pentatomoidea (Hemiptera-Heteroptera). Philippine Entomol. 8, 865 - 961.
Lent, H., Jurberg, J., 1965. Contribuic ¸ ao ao conhecimento dos Phloeidae Dallas, 1851, com um estudo sobre genitalia (Hemiptera, Pentatomoidea). Rev. Bras. Biol. 25, 123 - 144.
Leston, D., 1953 b. '' Phloeidae' ' Dallas: systematics and morphology, with remarks on the phylogeny of '' Pentatomoidea' ' Leach and upon the position of '' Serbana '' Distant (Hemiptera). Rev. Brasil. Biol. 13, 121 - 140.
Rider, D. A., 2006. Pentatomoidea Home Page. North Dakota State University. http: // www. ndsu. nodak. edu / ndsu / rider / Pentatomoidea / [accessed on 21 July 2006].
Rolston, L. H., McDonald, F. J. D., 1979. Keys and diagnoses for the families of Western Hemisphere Pentatomoidea, subfamilies of Pentatomidae and tribes of Pentatominae (Hemiptera). J. NY Entomol. Soc. 87, 189 - 207.
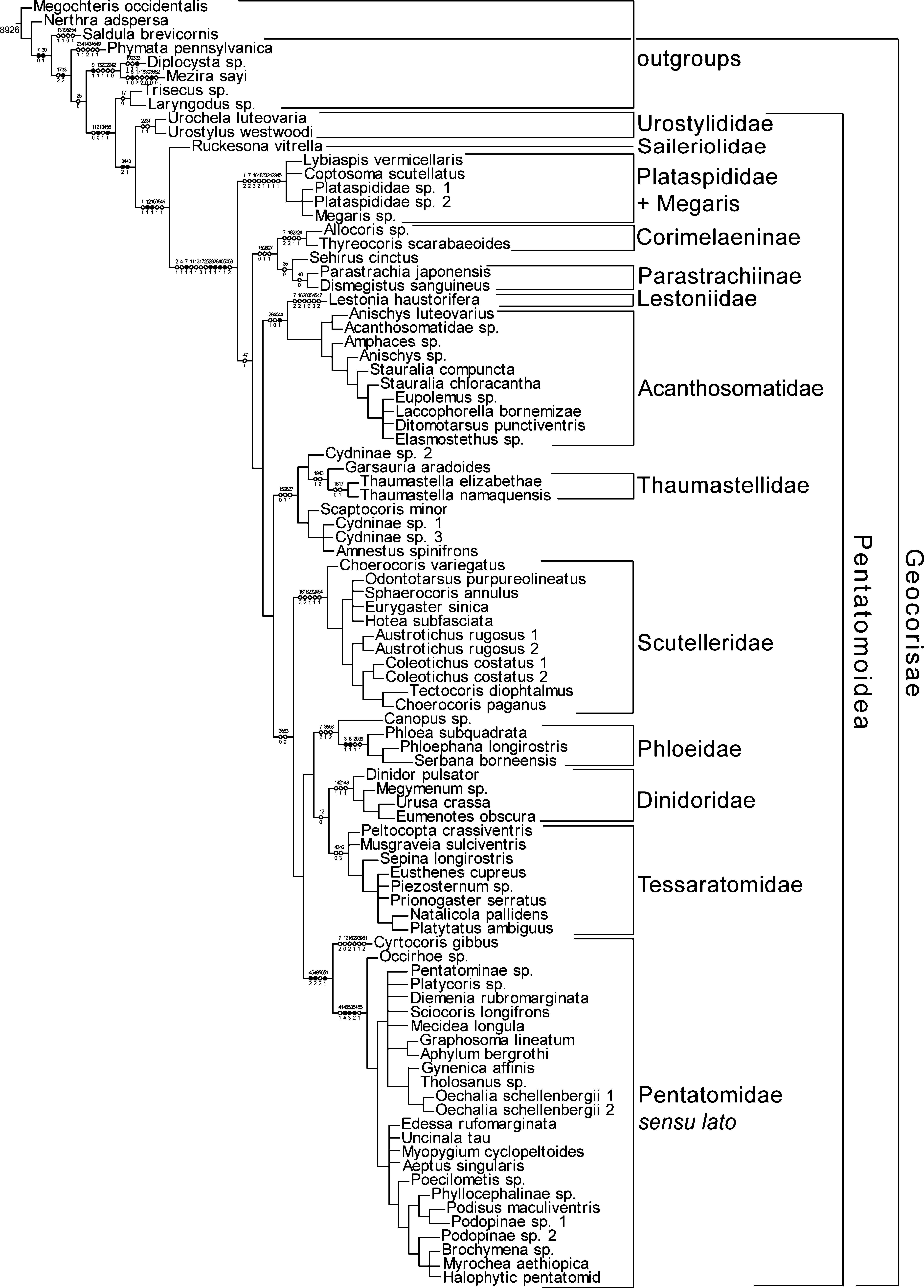
Fig. 54. One of four trees from total evidence analysis with POY of 92-taxon data set using 1: 2 indel ⁄ transition–transversion cost ratio. (d) Non-homoplasious; (s) homoplasious.

Fig. 55. One of three trees from total evidence analysis with POY of 92-taxon data set using 2: 2 indel ⁄transition–transversion cost ratio, which had the lowest MRI value. Bremer support values are shown.

Figs 45–48. 45. One of two trees derived from analysis of combined molecular data with 1: 1 indel ⁄transition–transversion cost ratio. 46. Single tree derived from analysis of combined molecular data with 2: 2 indel ⁄ transition–transversion cost ratio. 47. Single tree derived from analysis of ~500 bp of 16S rRNA data using 1: 1 indel ⁄ transition–transversion cost ratio. 48. Single tree derived from analysis of ~1800 bp of 18S rRNA using 1: 1 indel ⁄ transition–transversion cost ratio.
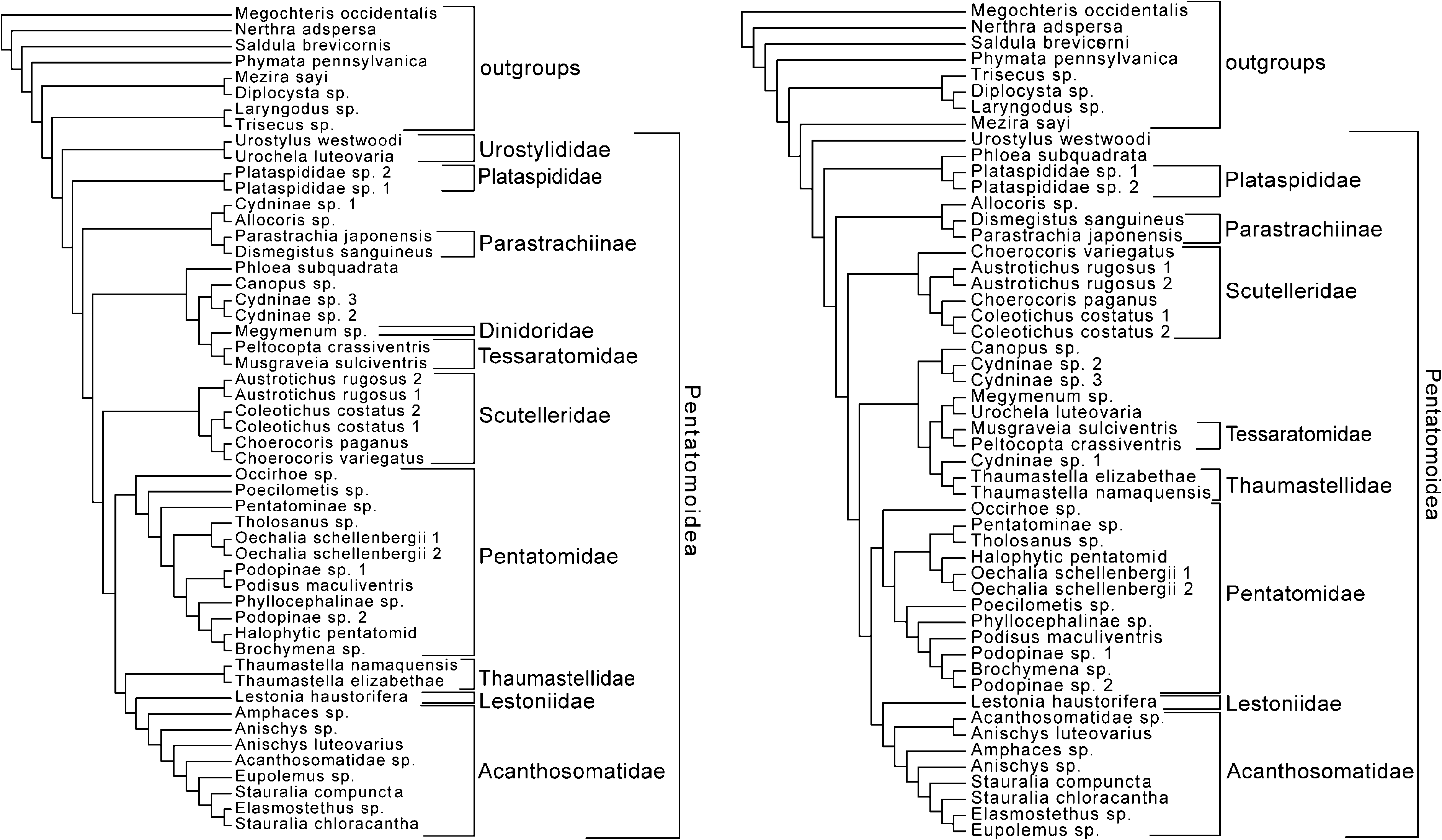
Figs 49–52. 49. Single tree derived from analysis of ~470 bp of 28S rRNA using 1: 1 indel ⁄ transition–transversion cost ratio. 50. Single tree derived from analysis of ~1100 bp of COI mtDNA using 1: 1 indel ⁄transition–transversion cost ratio. 51. Total evidence analysis with POY of 52- taxon data set using 1: 1 indel ⁄ transition–transversion cost ratio. 52. Total evidence analysis with POY of 52-taxon data set using 2: 2 indel ⁄ transition–transversion cost ratio.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Cimicomorpha |
|
SuperFamily |
Pentatomoidea |
|
Family |