
Metaleptobasis brysonima Williamson, 1915
|
publication ID |
https://doi.org/10.11646/zootaxa.3738.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:77D1A6F6-C320-442B-AF31-83324E5EAF3B |
|
persistent identifier |
https://treatment.plazi.org/id/03E187ED-662F-FFA4-D7A8-FE8CE748FC7C |
|
treatment provided by |
Felipe |
|
scientific name |
Metaleptobasis brysonima Williamson, 1915 |
| status |
|
Metaleptobasis brysonima Williamson, 1915 View in CoL
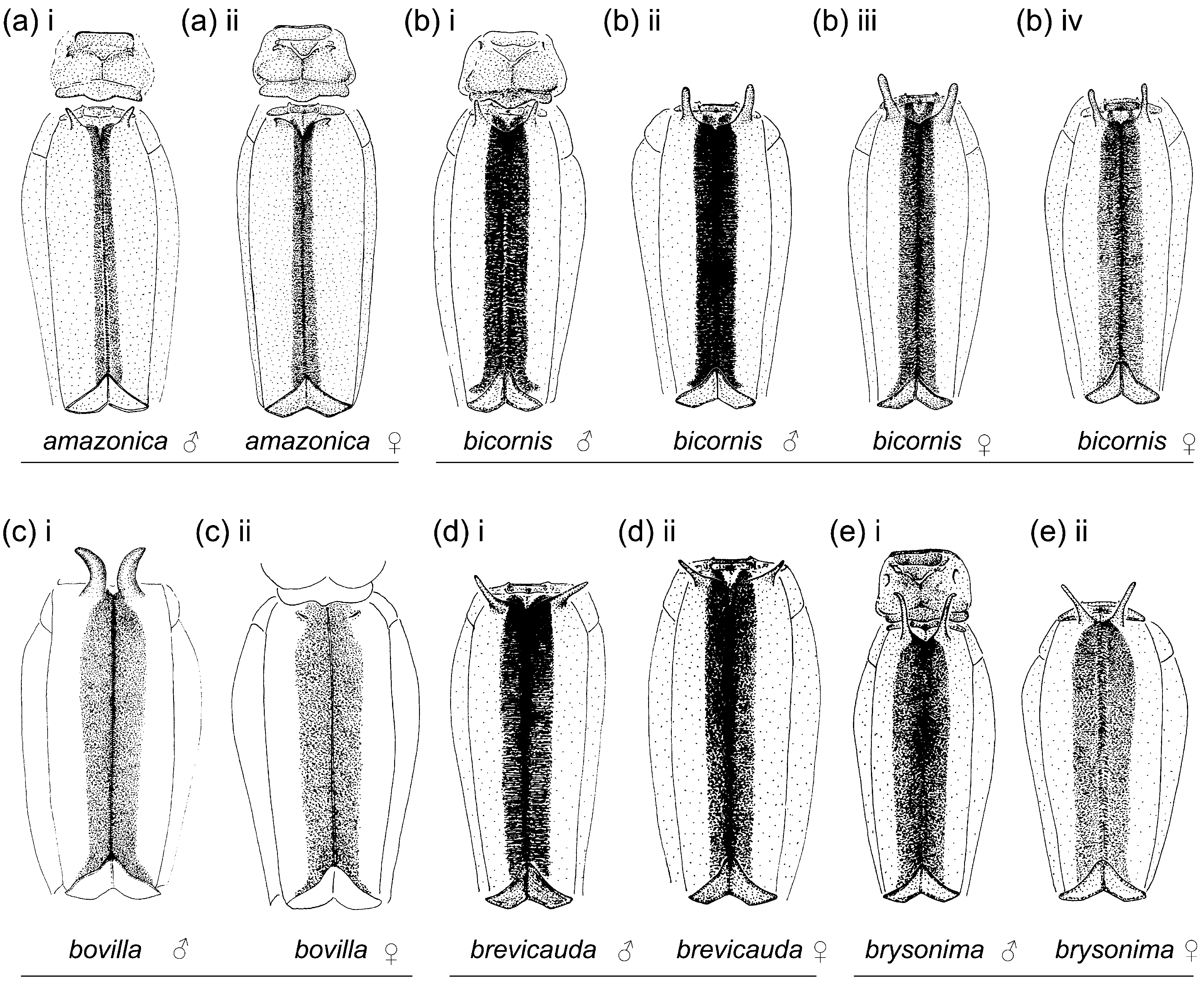
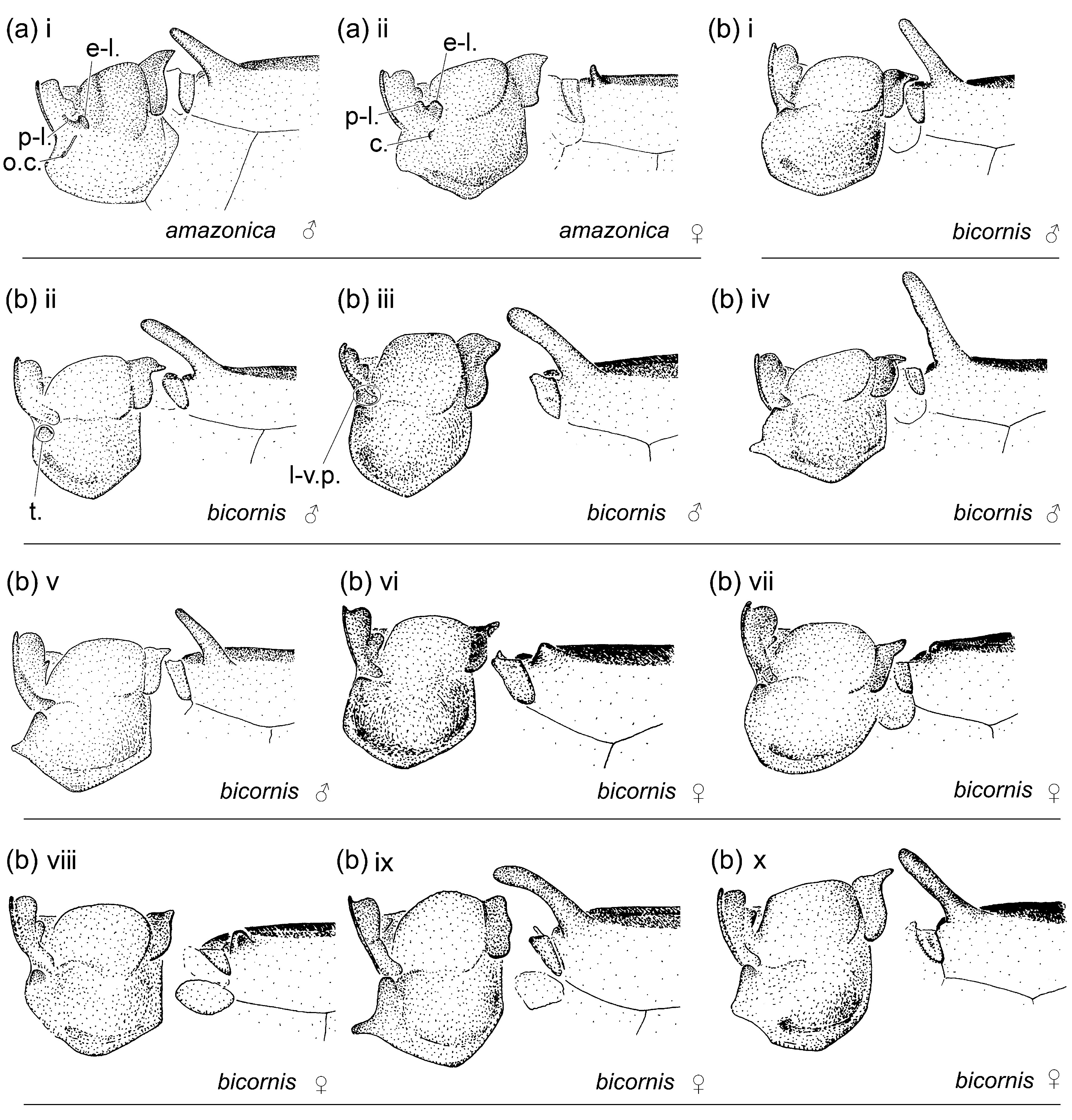
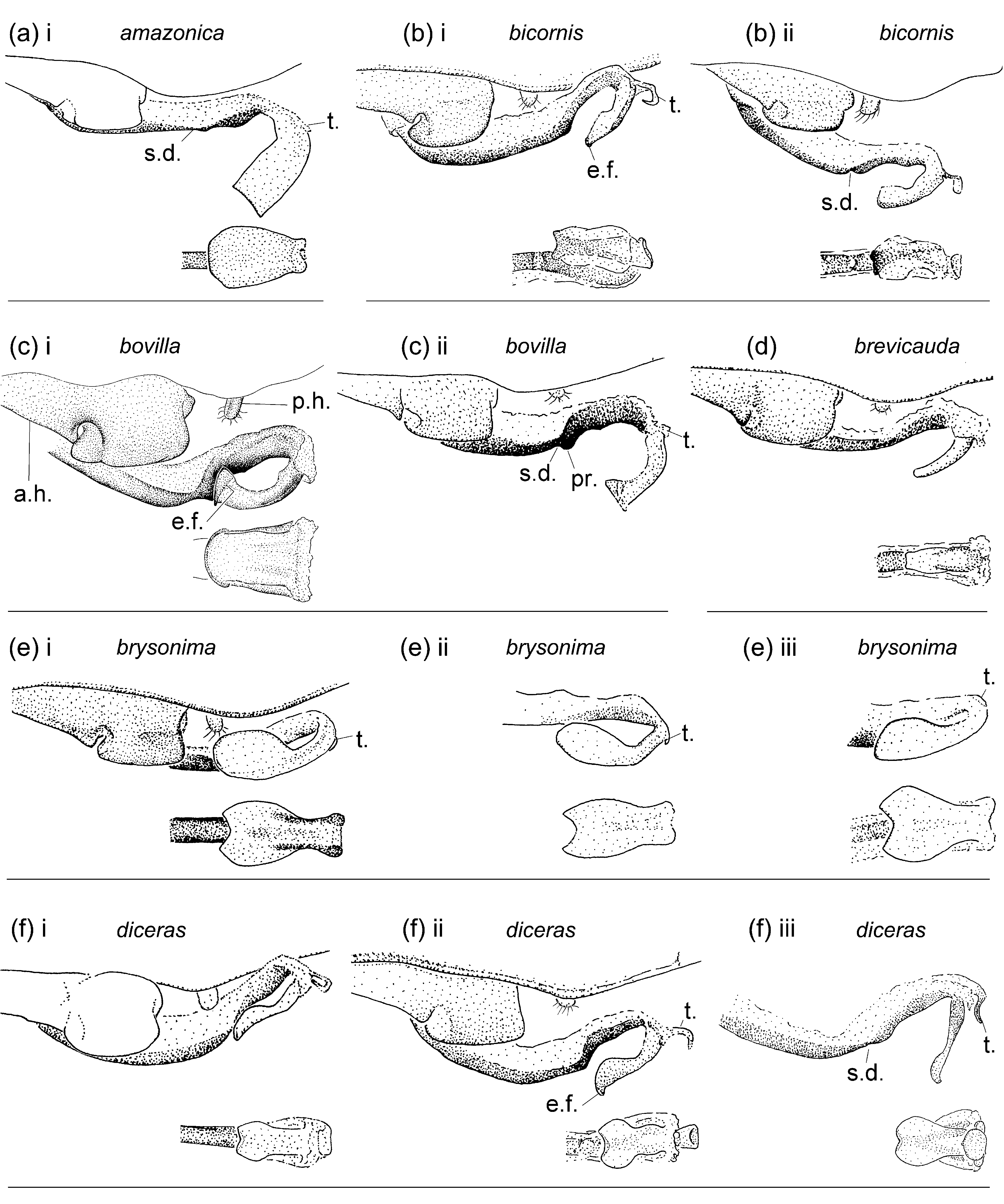
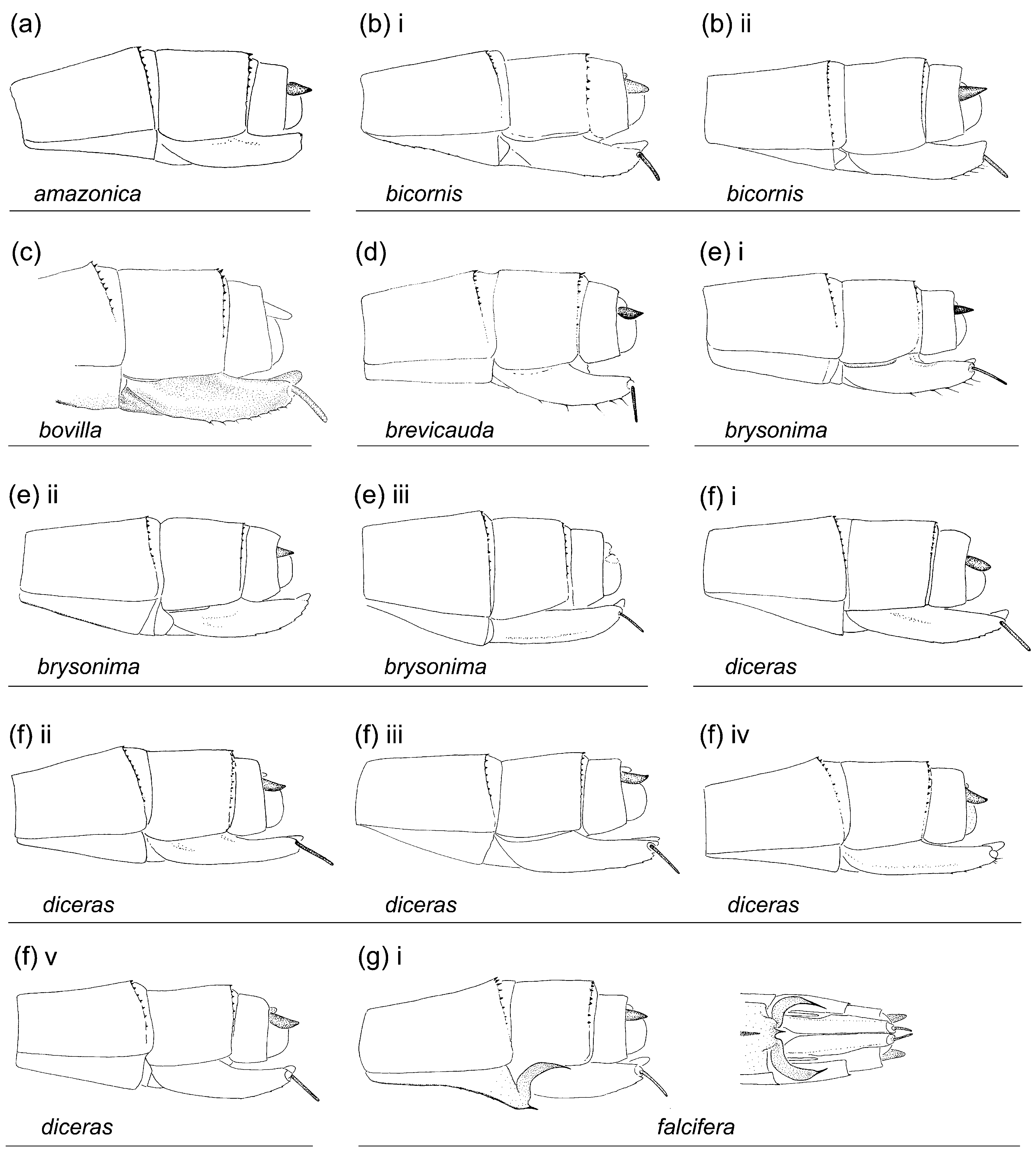
Figs. 1e View FIGURE 1 , 2b View FIGURE 2 , 3e View FIGURE 3 , 4e View FIGURE 4 , 5e View FIGURE 5 , 7 View FIGURE 7 , 8e View FIGURE 8 , 9e View FIGURE 9 , 10e View FIGURE 10 , 113, 12e, 13a, 14b
Metaleptobasis brysonima Williamson, 1915: 602 View in CoL , 604–607, figs. 5–8 (description of ♂, illustrations of S10 and mesanepisternal horns);— Geijskes (1932: 261; mention);— Cumming (1954: 29, 31; inclusion in key to males, proposed grouping);— Santos (1956: 384; comparison with M. selysi View in CoL );— Michalski (1988: 45–46; in part, mention from Trinidad, inclusion in key);— Daigle (2003: 373; comparison with M. minteri View in CoL );—Garrison et al. (2003: 27; inclusion in UMMZ type catalog, transcription of holotype labels);— Heckman (2008: 401, fig. 3.1.464; inclusion in key, illustration of wings, posterior lobe of pronotum, and mesanepisternal horns from Rácenis 1955);— Garrison et al. (2010: 284; mention).
Metaleptobasis byrsonima View in CoL — Williamson (1917: 8; unjustified emendation of name).
Metaleptobasis tetragena Calvert, 1945: 60–61 View in CoL , figs. 20–22 (description of ♀, illustrations of pronotal hind lobe and mesanepisternal horns);— Cumming (1954: 27, 30; mention);— Heckman (2008: 397; inclusion in key);— Garrison et al. (2010: 284; mention). New synonymy.
Metaleptobasis weibezahni Rácenis, 1955: 15–19 View in CoL , figs. 2c (description of ♀, illustrations of pronotal hind lobe and mesanepisternal horns);— De Marmels (1989: 32, figs. 124–126; comparison ♀, illustrations of pronotal hind lobe and mesanepisternal horns ♀ holotype);—De Marmels (1990: 337; mention);— Heckman (2008: 397–398, fig. 3.1.459; inclusion in key, illustration of posterior lobe of pronotum and mesanepisternal horns from De Marmels 1989);— Garrison & von Ellenrieder (2009: 48; mention);— Garrison et al. (2010: 284; mention). New synonymy.
Metaleptobasis sp. A — Paulson (1985: 12; mention).
Metaleptobasis incisula De Marmels 1989: 31–32 View in CoL , figs. 117–123 (description of ♂ ♀, illustrations of ♀ pronotal hind lobe and mesanepisternal horns, ♂ S10 and genital ligula);—De Marmels (1990: 337; mention);— De Marmels (1992a: 41; mention);— Heckman (2008: 398–399, fig. 3.1.461; inclusion in key, reproduction of illustration of posterior lobe of pronotum and mesanepisternal horns from De Marmels 1989);— Garrison & von Ellenrieder (2009: 48; mention);— Garrison et al. (2010: 284; mention). New synonymy.
Types. Holotype ♂ *: TRINIDAD, St. Andrew County, Cumuto , small swamp { 10°35'N, 61°12'W, 57 m}, 10 iii 1912, E.B. Williamson, L.A. Williamson & B.J. Rainey leg. [ UMMZ]. GoogleMaps
Specimens examined. Total: 26 ♂, 20 ♀: VENEZUELA, Delta Amacuro State : 1 ♀ holotype of M. weibezahni, Güiniquina { 9°11'N, 61°8'W, 15 m}, 10 viii 1954, Cermeli leg. [ MIZA]; Amazonas State GoogleMaps : 1 ♂, 1 ♀ paratypes of M. incisula, Atabapo, Sima Marawaka , macizo central { 3°34'N, 65°27'W, 1140 m}, 3–17 iii 1985, R. Candia & A. Chacón leg. [ FSCA] GoogleMaps ; 1 ♂ paratype of M. incisula , same but 3–10 iii 1985 [ USNM] ; 1 ♂ 1 ♀ paratypes of M. incisula , same but 10–17 iii 1985, Exp. Terramar [ RWG] ; 1 ♂, 1 ♀, Río Baria, Neblina Base Camp { 0°50'N, 66°9'W, 140 m}, 15 ii 1984, P.J. Spangler, P.M. Spangler & R.A. Faitoute [ USNM]; Bolívar State GoogleMaps : 3 ♂, wooded swamp 4 km W of El Paují, near Río Chaverú ( 4°28'N, 61°37'W, 1,000 m), 6 viii 1990, RWG leg. [ RWG] GoogleMaps ; 1 ♂, same but [ FSCA] ; 1 ♂, same but 900 m, TWD leg. [ FSCA]; GUYANA, Bartica Dis .: 1 ♀ holotype of M. tetragena, Kartabo { 6°22'N, 58°40'W}, vii 1926, C. Hodge IV leg. [ ANSP]; Cuyuni-Mazaruni Region GoogleMaps : 1 ♂, 1 ♀, Pakaraima Mountains near Mazaruni Headquarters { 6°24'N, 58°39'W, 0 m}, 1932, J.G. Myers leg. [ FSCA]; SURINAM, Sipaliwini Dis GoogleMaps .: 1 ♂, Wilhelmina Mountains , forest trail I, km 3–4, 17 viii 1943, D.C. Geijskes [ RMNH] ; 1 ♂, 1 ♀, same but sand creek, Kwatta camp N km 1, 3 ix 1943, D.C. Geijskes leg. [ RHMN] ; 1 ♂, Kappelsavanne , brown water forest creek { 3°48'N, 56°8'W, 280 m}, 14 iii 1958, D.C. Geijskes leg. [ RHMN] GoogleMaps ; 1 ♂, same but Kappelsavanne, forest trail S, near Mauritia palm forest, 20 iii 1958, D.C. Geijskes leg. [ RHMN] ; 1 ♂, same but Kappelsavanne , forest trail N, 24 iii 1958, D.C. Geijskes leg. [ RMNH] ; 1 ♀, Lucie River camp, swamp in forest near creek { 3°26'N, 57°13'W, 190 m}, 12 vii 1963, S. Ligorie leg. [ RMNH]; BRAZIL, Amazonas State GoogleMaps : 1 ♀ Presidente Figueiredo , Igarapé da Onça ( 2°0'52''S, 60°1'43''W), 16 iv 2008, P. Reis leg. [ INPA] GoogleMaps ; 1 ♀ Presidente Figueiredo, Est. de Balbina, km 24 { 2°1'13''S, 59°49'29''W}, 1–12 ix 2002, F. Fhilo, F.F. Barbosa, & U.C. Varredura leg. [ INPA]; Pará State GoogleMaps : 4 ♂, 1 ♀, Rio Xingu Camp, Altamira { 3°12'S, 52°11'W, 91m}, x 1963, R. von Diringshofen leg. [ MZUSP] GoogleMaps ; 2 ♀, same but xi 1963 [ MZUSP]; Rondônia State : 1 ♂, 1 ♀, Abuná { 9°42'S, 65°23'W, 112 m}, 21 iii 1922, J.H. Williamson & J.W. Strohm leg. [ USNM] GoogleMaps ; 1 ♂, 1 ♀, Porto Velho, Area Abunan, T 10 Aleatorio ( 8°46'S, 63°54'W, 86 m), 17 v 2010, Nogueira & Mendes leg. [ MZUSP]]; [ MZUSP]; BOLIVIA, Beni Dep GoogleMaps .: 1 ♂, 1 ♀, Río Benicito, Chacobo Indian Village , forest trail ( 12°20'S, 66°00'W), 1–31 vii 1960, B. Malkin leg. [ UMMZ]; PERU, Madre de Dios Dep GoogleMaps .: 3 ♂, 2 ♀, Explorer's Inn on Río Tambopata , 30 km SW Puerto Maldonado, forest trail, swamp, sandy stream, and muddy stream ( 12°50'12''S, 69°17'36''W, 300 m), 10–30 vii 2002, DRP & N. Smith leg. [ DRP] GoogleMaps ; 1 ♀, Tambopata Nature Reserve, Boca Río La Torre ( 12°50'18''S, 69°17'59''W, 300 m), 20 ii 1982, G. Lamas leg. [ DRP] GoogleMaps ; 1 ♂, same but Tambopata Nature Reserve , 30 km SW Puerto Maldonado, forest trail ( 12°30'S, 69°12'W, 300 m), 14 ii 1982, F. Estremadoyro leg. [ DRP] GoogleMaps ; 1 ♀, same but swamp forest, 29 iii 1982, M. Frisbie leg. [ DRP] .
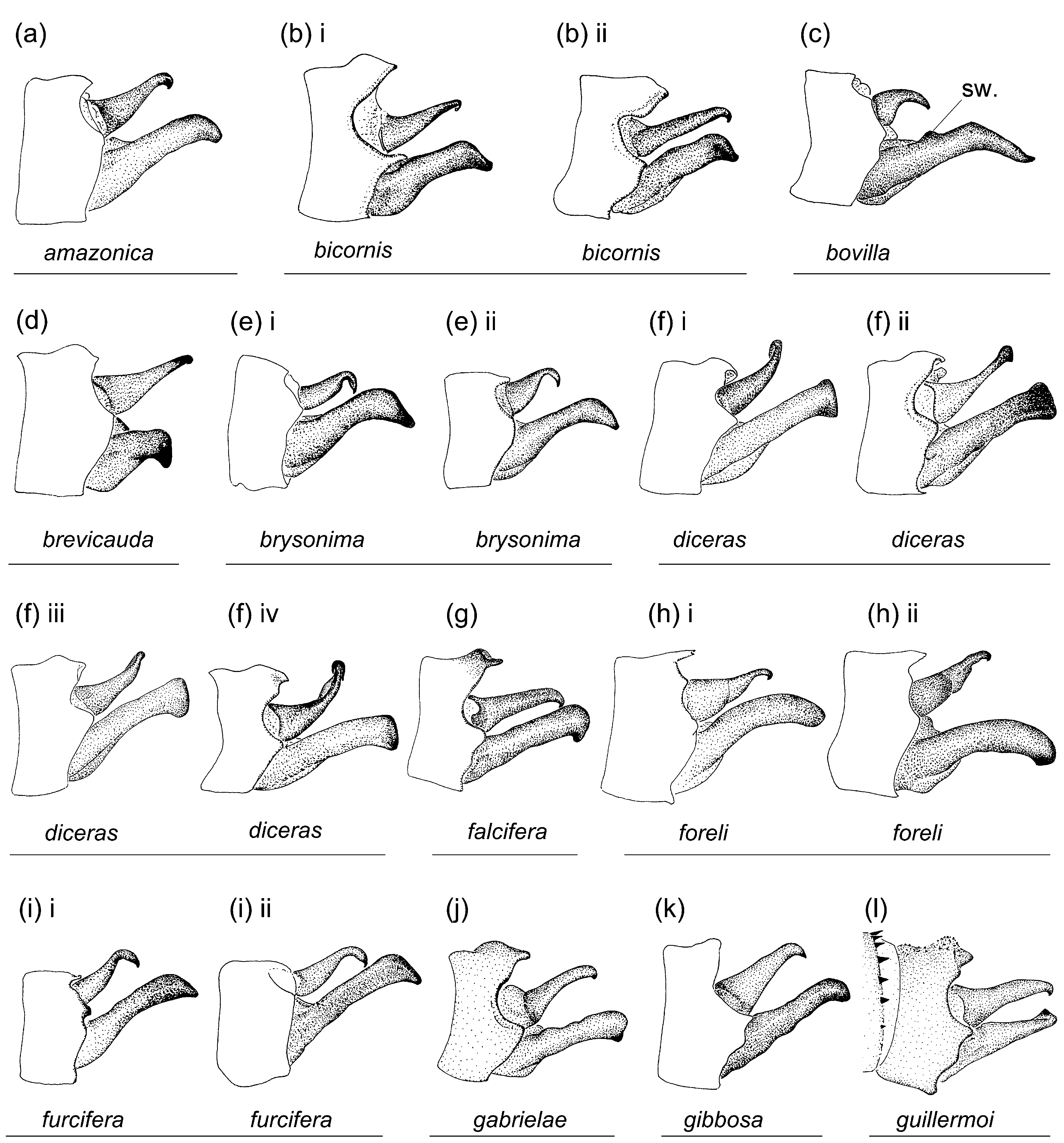
Characterization. Head: Labrum mostly pale; black on dorsum of head extensive; postocular lobes markedly angled ( Figs. 1e View FIGURE 1 ; 2b View FIGURE 2 ); eyes in life light green, or orange, or light green with dorsal black spot and orange medial third (DRP pers. comm.). Thorax: Pronotum anterior lobe ( Figs. 4e View FIGURE 4 ; 5e View FIGURE 5 ) smooth; anterior and middle lobes of pronotum separated dorso-laterally by a groove; anterior portion of propleuron with a low rounded tubercle (t.); a lateral c-shaped crest (c.) postero-dorsal to tubercle on propleuron; anterior margin of middle lobe of pronotum smooth; pronotum posterior lobe trilobed, with a central point on middle lobe in male, with a medial u- or vshaped incision in female, lateral margins rounded; mesanepisternal horns thin, with bases adjacent, in male as long as about two times mesostigmal plate width or slightly longer, oriented antero-dorsally and diverging only slightly except for pointed and laterally directed tips, forming an angle of 35°–45° with dorsum in lateral view ( Figs. 4e i View FIGURE 4 –iv), in female as long as 1.5–2 times mesostigmal plate width or slightly longer, directed antero-dorsally, with tips bluntly pointed, forming an angle of 45°–75° with dorsum in lateral view ( Figs. 4e v View FIGURE 4 –xiii); mid-dorsal dark stripe as wide as 0.50 of mesanepisterna, narrowing between mesanepisternal horns ( Figs. 2b View FIGURE 2 ; 3e View FIGURE 3 ); Pt subrectangular, with anterior and posterior sides longer than distal side. Abdomen: Male genital lobe short, less than 0.50 of anterior hamule height; posterior hamule digit-like and small, with at most only tip surpassing ventral margin of genital fossa in lateral view; curvature of basal segment of genital ligula marked by a slight concave depression; distal segment pear-shaped, with apex with a u-shaped incision surrounding lateral sides of basal segment at rest, lacking an ectal fold ( Figs. 7a i View FIGURE 7 ; 8e View FIGURE 8 ); posterior margin of female S8 sternum smooth, lacking any denticles, spines, or processes; distal end of ovipositor reaching apex of paraproct to apex of cercus ( Fig. 9e View FIGURE 9 ); medial portion of male S10 postero-dorsal margin ( Figs. 7a View FIGURE 7 ii–iii; 10e; 11e; 12e) projected posteriorly, with a small u-shaped incision, and lacking a dorsal prominence; male cercus about straight in dorsal view ( Fig. 10e View FIGURE 10 ), narrowing to tip, which forms a hook directed ventrally ( Figs. 7a View FIGURE 7 ii; 11e; 12e); ratio of male cercus length to S10 maximum length in lateral view 0.83–0.84; ratio of male cercus length to paraproct length in lateral view 0.47–0.53; male paraproct in lateral view narrowing distally, slightly wider at distal third than at medial third, with sides arched (7a ii; 12e); tip of paraproct with a faint ridge on medial surface ending on an apical tooth ( Figs. 10e View FIGURE 10 ; 11e View FIGURE 11 ).
Dimensions. Males ( n 10): Hw 20.7–23.4 [21.8 ± 1]; abdomen 33.4–40.6 [36.5 ± 2]; total length 40.3–48.1 [43.6 ± 2.2]. Females ( n 10): Hw 21.5–23 [22.3 ± 0.5]; abdomen 33–36.9 [35 ± 1.2]; total length 39.8–43.9 [41.9 ± 1.3].
Diagnosis. Metaleptobasis brysonima shares only with M. furcifera and M. paludicola the following combination of characters: small digit-like male posterior hamule, smooth female posterior margin of S8 sternum lacking any denticles, spines or processes, black on head dorsum covering about half to most of dorsal surface, labrum mostly pale, anterior margin of middle lobe of pronotum smooth, anterior portion of propleuron with a low rounded tubercle, and mesanepisternal horns sub-cylindrical. It differs from M. paludicola by female pronotum unlobed to slightly trilobed, always with a medial incision ( Figs. 4e v View FIGURE 4 –xiii; trilobed with smoothly convex medial lobe in M. paludicola , 4u v–vi), dark mid-dorsal pterothoracic stripe about as wide as 0.50 of mesanepisterna width ( Fig. 3e View FIGURE 3 ; about as wide as 0.33 of mesanepisterna width in M. paludicola , Fig. 3u View FIGURE 3 ), and male S 10 in lateral view flat, lacking a medial prominence on postero-dorsal margin ( Figs. 11e View FIGURE 11 ; 12e View FIGURE 12 ; with a well developed medial prominence on postero-dorsal margin in M. paludicola , Figs. 11u View FIGURE 11 ; 12u View FIGURE 12 ). It differs from M. furcifera by apex of male genital ligula distal segment concave, surrounding lateral sides of basal segment at rest ( Figs. 7a i View FIGURE 7 ; 8e View FIGURE 8 ; apex transverse and not extending over lateral sides of basal segment at rest in M. furcifera , Fig. 8i View FIGURE 8 ), and tip of male paraproct with a faint low longitudinal crest on inner surface ending on a single apical tooth pointed inwards ( Figs. 7a View FIGURE 7 iii; 10e; 11e; with a marked longitudinal crest on inner surface with two teeth pointed inwards, one on its basal end and one on its apical end in M. furcifera , Figs. 10i View FIGURE 10 ; 11i View FIGURE 11 ). In females, mesanepisternal horns are relatively shorter, 1.5–2 times as long as mesostigmal plate width, and slightly thicker in M. brysonima (in M. furcifera 2–3 times as long as mesostigmal plate width and slightly thinner), but I found no other structural or color characters to discriminate between females of these two species. Both species seem to be allopatric, with M. brysonima widely distributed from Trinidad, Venezuela, Guyana, N Brazil, to SE Peru and N Bolivia, and M. furcifera restricted to N Peru ( Fig. 14b). All known females of M. brysonima are dimorphic in the morphology of posterior lobe of pronotum and mesanepisternal horns, and both characters show a considerable degree of plasticity ( Figs. 4e v View FIGURE 4 –xiii; 5e iii–xi).
Remarks. Williamson (1915) described M. brysonima based on a single male from Trinidad. According to a footnote (page 602) in the description, the species was named after the genus of a small tree found in the savannah near the type locality, and the name was given to Williamson by Mr. W.E. Broadway from the Botanic Garden in Tobago. Later Williamson noted that the correct generic name of the plant was Byrsonima, not Brysonima, and he published a note proposing to change the spelling of the name to reflect the correct name of the tree ( Williamson 1917). However, according to the International Code of Zoological Nomenclature, the correct name must be the original spelling (Article 32.2), unless there is clear evidence of an inadvertent error and the corrigendum is issued simultaneously with the original work or as a circulated slip to be inserted in the work (or if in a journal, or work issued in parts, in one of the parts of the same volume) (Article 32.5.1). Since Williamson’s (1917) correction was published a posteriori in a different journal, his correction is an unjustified emendation, and the original spelling of the name has since been used by all subsequent authors.
Calvert (1945) described M. tetragena based on a single female from Guyana, and diagnosed it only from other known females. Association of males and females of M. brysonima , and examination of the holotype of M. tetragena , shows that this name represents a junior subjective synonym of M. brysonima . The holotype specimen of M. tetragena is a very teneral female, partially crushed, and with color pattern not fully developed. The pronotum is slightly compressed laterally, and the ‘two erect processes’ on the hind margin of pronotum that Calvert (1945) mentioned are in fact the sides of the medial cleft that characterizes the female pronotum in M. brysonima ( Figs. 4e v View FIGURE 4 –xiii; Figs. 5e View FIGURE 5 iii–xi), which are slightly twisted sideways ( Figs. 4e View FIGURE 4 ix; 5e vii).
Based on similarity of a male collected in Venezuela to the description of the holotype of M. brysonima, Rácenis (1955) described co-occuring females as representing this species. He did not examine type material and instead based his identifications on original descriptions. The original descriptions of M. brysonima and M. quadricornis are brief and in the case of M. quadricornis are also open to misinterpretation due to lack of illustrations. Since male cerci and color pattern in M. brysonima and M. quadricornis are very similar, unfortunately he misassociated these two names. After examining the holotype female of M. quadricornis and the holotype male of M. brysonima , as well as pairs of males and females of both species, I was able to conclude that the species Rácenis (1955) identified as M. brysonima and for which he described and illustrated the female (figs. 2d, mislabeled as 2c as stated by De Marmels 1989: 32) is in fact M. quadricornis . Even though the male of M. brysonima resembles that of M. quadricornis by cercus shape and general color pattern, M. brysonima can be distinguished from M. quadricornis by anterior margin of middle lobe of pronotum smooth ( Figs. 4e View FIGURE 4 ; 5e View FIGURE 5 ; forming a dorsal crest followed by a transverse depression in M. quadricornis , Figs. 4y View FIGURE 4 ; 5y View FIGURE 5 ), posterior lobe of female pronotum unlobed to slightly trilobed always with a medial u to v-shaped incision ( Figs. 4e v View FIGURE 4 –xiii; trilobed with lateral lobes rounded and surpassing medial lobe posteriorly in M. quadricornis , Figs. 4y View FIGURE 4 iii–vii), and male mesanepisternal horns directed anteriorly at an angle of 35°–45° with dorsum ( Fig. 5e i View FIGURE 5 –ii; directed dorso-anteriorly at 60°–85° with dorsum in M. quadricornis , Fig. 5y i View FIGURE 5 –iii).
In the same paper, Rácenis (1955) described a single female, representing a different species from M. brysonima sensu Rácenis (= M. quadricornis ), as M. weibezahni (figs. 2c, mislabeled as 2d as stated by De Marmels 1989: 32). Examination of the holotype female of M. weibezahni ( Figs. 1e View FIGURE 1 iii; 3e iii; 4e viii; 5e vi; 9e ii) and knowledge of the range of variability of the characters of the females of M. brysonima , allows me to conclude that this female represents a specimen of M. brysonima , and that M. weibezahni is consequently a junior subjective synonym of M. brysonima .
De Marmels (1989, 1992a, b) followed Rácenis’ (1955) species treatment that considered M. quadricornis to represent M. brysonima and, based on several males and females of M. brysonima , described M. incisula as a new species ( De Marmels 1989). Comparison of his description and illustrations (figs. 117–123) and of several paratypes of M. incisula ( Figs. 1e i View FIGURE 1 –ii; 3e i–ii; 4e ii, v–vi; 5e i, iii–iv; 8e i; 12e i) with the holotype male of M. brysonima ( Figs. 2b View FIGURE 2 ; 6 View FIGURE 6 ) and numerous additional males and females from several localities allows me to conclude that M. incisula is a subjective junior synonym of M. brysonima . De Marmels (1989: 31) suggested that M. incisula (= M. brysonima ) was perhaps allied to M. quadricornis , M. tetragena , and M. weibezahni but could be separated from them by its thoracic armature. As observed by De Marmels (1989) in his type series of M. incisula , I found that the medial incision characterizing the hind lobe of the pronotum of the female of M. brysonima can be of variable width and shape ( Figs. 4e v View FIGURE 4 –xiii) and, likely due to postmortem preservation, its sides can also be oriented not only posteriorly but also medially, dorso-posteriorly, or dorsally, as is the case in the holotypes of M. tetragena and M. weibezahni . Female mesanepisternal horns are always adjacent at base and diverge latero-dorsally to their apices in M. brysonima , but their exact length, width, orientation, and angle with dorsum of pterothorax are somewhat variable, as I also observed for mesanepisternal horns in other congeneric species for which large series of specimens are available (i.e. M. bicornis , M. foreli , M. lillianae , M. minteri , and M. quadricornis ). The shorter horns of the female holotype of M. tetragena and the more upright horns of the female holotype of M. weibezahni both fit within the range of variability I observed for females of M. brysonima ( Figs. 4e v View FIGURE 4 –xiii; 5e iii–xi).
Based on literature records, Michalski (1988: 45) listed M. brysonima from Trinidad and included a key for the three species of Metaleptobasis known from that island; the characters he used in the key will correctly key out males from Trinidad, but his female characters—probably taken from Rácenis (1955) —correspond to females of M. quadricornis .
Machet (1989: 14) recorded M. brysonima from French Guiana. Unfortunately, I was unable to examine a specimen to determine whether it was really M. brysonima or rather M. quadricornis if he followed Rácenis’ (1955) treatment.
Habitat. Forest near rivers, swamps, creeks, and small streams. Dennis Paulson observed several specimens in Tambopata, Peru, perching low in the forest, from about 0.6 m to head height, along trails at swamps and near streams, and one male flying low at about 1.3 m above ground, not in bright sun ( DRP pers. comm.) .
Distribution. Trinidad and Venezuela south to Guyana, Suriname, N Brazil, and SE Peru ( Fig. 14b).







| UMMZ |
University of Michigan, Museum of Zoology |
| MIZA |
Museo del Instituto de Zoologia Agricola Francisco Fernandez Yepez |
| FSCA |
Florida State Collection of Arthropods, The Museum of Entomology |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| ANSP |
Academy of Natural Sciences of Philadelphia |
| RMNH |
National Museum of Natural History, Naturalis |
| INPA |
Instituto Nacional de Pesquisas da Amazonia |
| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Metaleptobasis brysonima Williamson, 1915
| Ellenrieder, Natalia Von 2013 |
Metaleptobasis incisula
| Garrison, R. W. & von Ellenrieder, N. & Louton, J. A. 2010: 284 |
| Garrison, R. W. & von Ellenrieder, N. 2009: 48 |
| Heckman, C. W. 2008: 398 |
| De Marmels, J. 1992: 41 |
| De Marmels, J. 1989: 32 |
Metaleptobasis sp. A
| Paulson, D. R. 1985: 12 |
Metaleptobasis weibezahni Rácenis, 1955: 15–19
| Garrison, R. W. & von Ellenrieder, N. & Louton, J. A. 2010: 284 |
| Garrison, R. W. & von Ellenrieder, N. 2009: 48 |
| Heckman, C. W. 2008: 397 |
| De Marmels, J. 1989: 32 |
| Racenis, J. 1955: 19 |
Metaleptobasis tetragena
| Garrison, R. W. & von Ellenrieder, N. & Louton, J. A. 2010: 284 |
| Heckman, C. W. 2008: 397 |
| Cumming, R. B. 1954: 27 |
Metaleptobasis byrsonima
| Williamson, E. B. 1917: 8 |
Metaleptobasis brysonima
| Garrison, R. W. & von Ellenrieder, N. & Louton, J. A. 2010: 284 |
| Heckman, C. W. 2008: 401 |
| Daigle, J. J. 2003: 373 |
| Michalski, J. 1988: 45 |
| Santos, N. D. 1956: 384 |
| Cumming, R. B. 1954: 29 |
| Geijskes, D. C. 1932: 261 |
| Williamson, E. B. 1915: 602 |