
Aplysina fistularis (Pallas, 1766)
|
publication ID |
https://doi.org/10.5281/zenodo.178878 |
|
DOI |
https://doi.org/10.5281/zenodo.6240781 |
|
persistent identifier |
https://treatment.plazi.org/id/03E387FE-FFCD-FFED-E3B5-FDEEFB57BEA3 |
|
treatment provided by |
Plazi |
|
scientific name |
Aplysina fistularis (Pallas, 1766) |
| status |
|
Aplysina fistularis (Pallas, 1766) ( Figs. 3 View FIGURE 3 C, 5–6, Tab. III)
Spongia fistularis, Pallas (1766: 385) . Aplysina fistularis sensu Lendenfeld (1889: 422) . Verongia fistularis sensu de Laubenfels (1956: 2), Hechtel (1976: 239). Verongia fistularis forma fistularis sensu Collete & Rützler (1977: 309) . Further synonymy cf. Hechtel (1965: 12) and Wiedenmayer (1977: 64).
Type specimens status: All type material of Pallas was destroyed by fire in Küstrin ( Poland) in 1758 during the Seven Years' War, including the holotypes of A. fistularis and A. fulva ( Wiedenmayer, 1977) . It is likely that Pallas had actual specimens at hand because he mentioned (cf. Wiedenmayer, 1977) that A. fistularis (as Spongia fistularis ) was denser and harder than A. fulva (as S. fulva ). De Laubenfels’ (1948) designation of a neotype from Topsent’s (1931) redescription of Lamarck’s (1814) Spongia fistularis was considered unhappy by Wiedenmayer (1977), who argued that these specimens were far from representing the typical form of the species, where the specimen was a single large tube with brown colour when dry (cf. Seba, 1758; Esper, 1791 –1799; Ehlers, 1870). Additionally, he pointed out that de Laubenfels did not select which from three hypotypes in Lamarck´s collection he meant. We had the opportunity to reexamine the three hypotypes [ MNHN LBIM No D.T. 3379, plus another two unregistered specimens of Spongia fistularis var. aggregata ]. In spite of their cospecificity with Aplysina fistularis not being 100% ruled out, they are considerably atypical by consisting of clusters of relatively small, largely anastomosed tubes.
From repeatedly reading the above quoted references, it appears to us this whole argument is basically flawed, because it seems quite unlike that de Laubenfels might have overlooked Topsent’s clear mention of the existence of three specimens in Lamarck’s series of Spongia fistularis , referred by Topsent to Aplysina fistularis var. aggregata (= Aplysina insularis of authors). Rather, we prefer to consider that he was actually referring to Topsent’s redescription of Spongia tubaeformis Lamarck , referred by the former to Aplysina fistularis , and obviously comprising a single, illustrated specimen, much more likely to match the current concept of A. fistularis (e.g. van Soest, 1978; Zea, 1987; present description). This specimen is still extant in the MNHN Lamarck collection, registered under MNHN LBIM No D.T. 598, but reported to be broken at mid length in two equal pieces (I. Domart-Coulon, pers. comm.). Despite the fact that this description starts at page 71, instead of 72, as quoted by de Laubenfels (1948), it does continue into the latter page, where the specimen under consideration is actually described. If this argument is accepted, then de Laubenfels’ (1948) neotype designation fulfills a significant set of the qualifying conditions listed in ICZN’s Article 75 for recognition of the validity of any neotype designation. Nevertheless, it appears to us that conditions 75.3.4 and 75.3.6 were not satisfactorily matched. As recommended by the ICZN, a subsequent reviewer should prioritize the validation of a former (prior to 1961) invalid designation, which is attempted here. Condition 75.3.4 can be satisfied by reference to Wiedenmayer’s (1977) quotation of the Seven Years’ War fire at Küstrin which destroyed Pallas’ collection. The choice for Lamarck’s (1814) specimen was a natural one, as this possibly was the oldest available, reasonably described and illustrated specimen. Condition 75.3.6 is matched by the realization that Puerto Rico, type locality for Lamarck’s specimen falls within Pallas’ “Mare Americanum”. Unfortunately, the possibility that A. fistularis ’ neotype might belong to A. archeri (Higgin, 1875) instead cannot be entirely ruled out, because the live colour of Lamarck’s specimen was unknown.
Studied material: MNRJ 690, Sueste Island, east side (Parque Nacional Marinho dos Abrolhos, BA), C.A. Secchin coll., 16/IV/1981. MNRJ 1548, Pirapama´s shipwreck (off Recife, PE), G. Muricy coll., 20 m depth, 11/II/1998. MNRJ 2526, shipwreck, off Cantagalo Beach ( Salvador, BA, 12º56.211' S - 38º30.698' W), E. Hajdu coll., 5 m. depth, 03/VIII/1999. MNRJ 2666, Northern Breakwater, Todos os Santos Bay ( Salvador, BA, 12º57.7' S - 38º31.1' W or 12º58.3' S - 38º31.2' W), 5m depth, E. Hajdu coll., 01/VIII/1999. MNRJ 3523 (Reserva Extrativista de Corumbau, Prado, BA), G. Muricy coll., 14/XI/1999. MNRJ 4164, 4166, 4172, 4174, Salvador ´s Yacht Club ( Salvador, BA), E. Hajdu coll., 3–5 m depth, 4/VII/2001. MNRJ 5275, Salvador ´s Yacht Club ( Salvador, BA) U.S. Pinheiro coll., 3 m depth, 29/I/2002. MNRJ 5468, Parcel Paredes, south side, (Parque Nacional Marinho dos Abrolhos, BA, 17º56.800´S - 38º54.260' W), S. Ribeiro coll., 15 m depth, 01/ III/2002. MNRJ 5470, Patch Reef between Rosalina ´s shipwreck and Santa Bárbara Island (Parque Nacional Marinho dos Abrolhos, BA, 17º57.765' S - 38º40.271' W), U.S. Pinheiro and E. L. Esteves coll., 15 m depth, 4/III/2002. MNRJ 5471, Patch Reef between Rosalina ´s shipwreck and Santa Barbara Island, (Parque Nacional Marinho dos Abrolhos, BA, 17º57.750' S - 38º40.710' W), G. Muricy and E. Vilanova coll., 14 m depth, 4/ III/2002. MNRJ 5944, Patch Reef between Rosalina ´s shipwreck and Santa Bárbara Island (Parque Nacional Marinho dos Abrolhos, BA, 17º57.765' S - 38º40.271' W), U.S. Pinheiro and E. L. Esteves coll., 15 m depth, 4/III/2002.
Comparative material: Fragment from IdO-401 ( holotype of Aplysina ocracea ).
Diagnosis: Specimens consist of tubes which can be cylindrical, fusiform or slightly barrel-shaped, single or grouped, and topped by a large apical pseudoscule. Tubes can be as tall as 1 m, or more. The surface can vary from nearly smooth to very irregular with variably shaped projections, such as shallow-ridges, folds, digitiform processes and lobes. Shorter digitiform projections, frequently constricted at the base, the size of small fistules, can also be abundant. Specimens turn nearly black upon fixation.
Description of the species:
Shape: Individuals consist of tubes which can be stout-cylindrical, fusiform or slightly barrel-shaped, single or grouped, and topped by a large apical pseudoscule. Tubes can be as tall as 1 m, or more (up to 60 cm high and 9 cm wide on collected specimens). Smaller, presumably young individuals can be only 3 cm high and 0.8 cm wide. Tube walls vary from 0.5 to over 2 cm in thickness. These tubes can be smooth, finely conulose or very irregular, and many bear lateral projections (6A–E), which vary from shallow-ridges and folds, to digitiform processes and lobes. All the tubes possess large apical pseudoscula 0.8–7.5 cm in diameter ( Fig. 6B View FIGURE 6 , E), but small lateral oscula can be observed. In MNRJ 4164 and 4174 these pseudoscula have iris-type diaphragms, mainly in smaller tubes. These structures are commonly seen on live specimens, but easily lost on preserved material. In MNRJ 5471 and 5944, small digitiform projections (up to 1 cm high) rising from the large apical pseudoscula can be noticed. Live-colour is normally yellow or ochre-yellow (brownish). After preservation in ethanol, all specimens turned into purple, dark brown or black. The specimens are mostly very soft, apart from MNRJ 5471 which is very hard.
Skeleton: Choanosome formed by an irregular polygonal reticulation of spongin fibers, 37–275 Μm thick (average 92 Μm; Fig. 6D View FIGURE 6 ). They have an amber coloured bark, with the exception of specimen MNRJ 5471 which is reddish coloured. The pith can be black or amber, with thickness of 10–60 Μm (average 26 Μm; Fig. 6 View FIGURE 6 E).
TABLE III: Spongin fibres’ measurement data for Aplysina fistularis (Pallas, 1766) (in micrometers; S.D. = Standard Deviation and N=30).
Specimens Locality* Fibers Piths
Thinnest Mean Thickest S.D. Thinnest Mean Thickest S.D. *BA, Bahia State; PE, Pernambuco State.
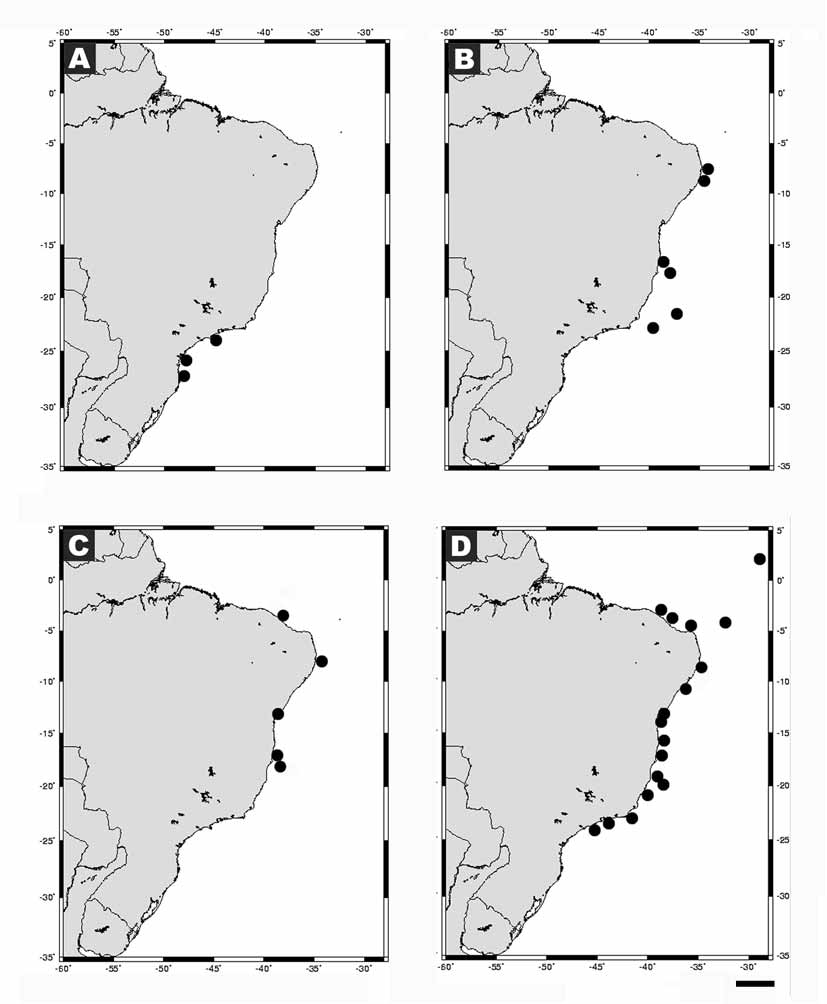
Distribution: Tropical western Atlantic: Brazil (3–18º S, Fig. 3 View FIGURE 3 C): Ceará State, Pernambuco State, Bahia State, Parque Nacional Marinho dos Abrolhos. World: British Virgin Islands, Cuba, Cozumel, San Andres, Turks & Caicos, Curaçao.
Ecology: The specimens were collected in warm and clear waters; smaller specimens were found as shallow as 4 m deep, larger ones deeper than 12 m. Aplysina fistularis is one of the most conspicuous and abundant sponges of Parque Estadual da Pedra da Risca do Meio (off Fortaleza, Ceará).
Remarks: Alcolado (1984) described Aplysina ocracea from Cuba mentioning that it differed from A. fistularis by the possession of thicker fibers and the preservation of its natural colouration (ochre) after fixation. However, in the A. fistularis material examined here, we can observe a large variation of fiber thickness that includes the variation described for A. ocracea (small fragment of the holotype, IdO-401, has been reexamined). A study of morphological variability of whole specimens is necessary to settle its limits as regards the variability of A. fistularis . For the present, we take P. Alcolado´s (pers. comm.) that the species is pedunculate as evidence that it deserves the status of a valid species.
Aplysina fistularis is a considerably plastic species, which stretches a continuum between small, smooth tubes; to very large ones, full of variably shaped projections, with or without their own apical pseudoscule. Such smaller, apparently solid projections were already reported by de Laubenfels (1948) as partially bridging the gap between A. fistularis and A. fulva . Other species described here may approach the limits of this variability, but we consider that morphologic gaps exist, and these will be discussed below.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Aplysina fistularis (Pallas, 1766)
| Pinheiro, Ulisses Dos S., Hajdu, Eduardo & Custódio, Márcio R. 2007 |
Spongia fistularis
| Collete 1977: 309 |
| Wiedenmayer 1977: 64 |
| Hechtel 1976: 239 |
| Laubenfels 1956: 2 |
| Lendenfeld 1889: 422 |