
Rineloricaria langei, Ingenito & Ghazzi & Duboc & Abilhoa, 2008
|
publication ID |
https://doi.org/10.1590/S1679-62252008000300009 |
|
DOI |
https://doi.org/10.5281/zenodo.17106719 |
|
persistent identifier |
https://treatment.plazi.org/id/03E687BD-FF99-FFC3-FC56-8DEB9858FBB6 |
|
treatment provided by |
Carolina |
|
scientific name |
Rineloricaria langei |
| status |
sp. nov. |
Rineloricaria langei View in CoL , new species
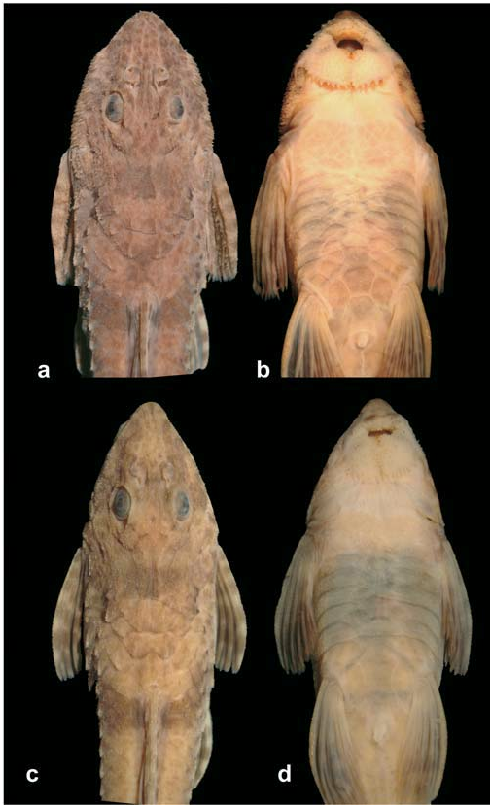
Rineloricaria sp. ; Ingenito et al., 2004: key, picture and comments (in part, only MHNCI 8937). [Picture shows a specimen from MHNCI 8937, but legend erroneously indicates MHNCI 9133].
Holotype. MCP 42506 , male, 114.9 mm SL, Brazil, Paraná, Quatro Barras, rio Iraí , at Fazenda Moura-Daz , downstream Reservoir of rio Iraí – SANEPAR, 49°05’26”W 25°29’23”S, Jan 2003, V. Abilhoa. GoogleMaps
Paratypes. MCP 23459 , 4 ( 1 male, 3 females), 71.7-116.5 mm SL GoogleMaps . MHNCI 8937 View Materials , 10 ( 1 male, 9 females), 51.7-111.9 mm SL GoogleMaps . MNRJ 31157 View Materials , 5 ( females, 1 c&s), 81.1-106.2 mm SL. All specimens collected with the holotype GoogleMaps .
Diagnosis. Rineloricaria langei can be distinguished from all species of the genus except R. catamarcensis , R. longicauda , R. maquinensis , R. misionera , R. nigricauda , R. quadrensis , and R. steindachneri by its narrow body (cleithral width 13.0-15.7% of SL; vs. 17.9-22.4% in R. aequalicuspis , 20.0-22.2% in R. anhaguapitan , 17.8-19.2% in R. anitae , 18.0- 21.8% in R. baliola , 16.0-19.1% in R. cadeae , 18.3-22.6% in R. capitonia , 17.7% in R. felipponei , 21.7-22.0% in R. jaraguensis , 17.1-20.6% in R. kronei , 19.1-20.6% in R. latirostris , 16.8-19.3% in R. malabarbai , 18.3-24.0% in R. microlepidogaster , 17.9% in R. pareiacantha , 18.7-19.9% in R. pentamaculata , 18.3-23.6% in R. reisi , 17.6-19.6% in R. tropeira , 16.0-18.1% in R. sanga , 17.6-19.9% in R. setepovos , 16.6-19.9% in R. stellata , 15.8-21.4% in R. strigilata , 18.2% in R. thrissoceps , and 16.2-21.6% in R. zaina ). Rineloricaria langei differs from R. catamarcensis , R. nigricauda and R. steindachneri by having the dorsal caudal ray not prolonged as a filament ( vs. dorsal caudal ray produced as a short filament). It is distinguished from R. quadrensis by having larger interorbital width (19.7-24.0% vs. 18.5-19.3% of HL) and longer dorsal-spine length (18.3-22.2% vs. 17.4-18.6% of SL); differs from R. longicauda by elliptical naked area at tip of snout, not reaching most anterior pore of the infraorbital sensory canal ( vs. elongate naked area at tip of snout reaching the most anterior pore of the infraorbital sensory canal). In addition, R. langei can be easily distinguished from R. maquinensis (with abdomen almost naked) and R. misionera (with pectoral girdle not covered by plates) by abdomen and pectoral girdle completely covered by plates.
Description. Morphometric data in Table 1 View Table 1 . Head and body deeply depressed. Head narrow in females and immature males, and broad in mature males. Body narrow. Body depth greatest at dorsal-fin origin. Head triangular in dorsal view. Dorsal profile of head convex from snout tip to anterior margin of parietosupraocciptal, thereafter slightly concave to posterior margin of first predorsal plate. Predorsal region straight. Trunk almost straight and gradually depressed from dorsal-fin origin to caudal-fin base. Ventral profile of body nearly straight from snout tip to anal-fin origin, then becoming more depressed towards caudal-fin base. Caudal peduncle depressed forming lateral keels with 12(1), 13(9), 14(8*), or 15 (2) coalesced plates from 29(6*), 30(13) or 31(1) total lateral plates in middle series. Four lateral series of plates, mid-dorsal series absent.Abdomen and ventral region of pectoral girdle completely covered by plates, even in specimens smaller than 85.5 mm SL; plates almost reaching transverse line crossing through anterior margin of branchial openings. Ventral region of scapular girdle covered by several irregularly arranged platelets smaller than abdominal plates. One (2), two (1), three (16*) or four (1) series of abdominal plates, usually regularly distributed in adults. Abdominal plates of irregular shapes and sizes. Pre-anal plate present and anteriorly rounded by three polygonal plates. Lateral abdominal plates four (1), five (9*), six (9) or seven (1). Dorsal-fin base with five plate rows. Anal-fin base with two (8) or three (12*) plate rows. Two (9) or three (11*) rows of plates between urogenital pore and anal fin. Plate counts on opposite sides of body usually different, except at dorsal- and anal-fin bases.
Top of head and parieto-supraocciptal wrinkled, with welldeveloped ridges. Ridges of parieto-supraocciptal diverging posteriorly in adult specimens and nearly parallel in young. Predorsal ridges parallel. Plates of first three mid-dorsal series with evident ridges. Upper edge of orbit raised. Postorbital notch shallow, short and wide, not surpassing one third of orbital diameter.
Head and body covered by small odontodes, somewhat more developed on top of head. Mature males with hypertrophied odontodes distributed only on lateral margins of head and dorsum of pectoral fin ( Fig. 2 View Fig ). Snout tip with small elliptical naked area, not reaching most anterior pore of infraorbital ramus of sensory canal. Naked area of snout separated from upper lip by two to four series of inconspicuous odontodes. Lips well developed and covered by papillae; only one irregular row of papillae at anterior most area of upper lip. Two rows of inconspicuous papillae separating upper and lower lips. Maxillary barbel thin and shorter than orbital diameter. Notch present in lower lip. Teeth bicuspid with lateral cusp smaller than medial. Premaxilla with five (3), six (8), seven (7) or eight (2*) teeth. Dentary with five (2), six (6), seven (6) or eight (6*) teeth. Total number of vertebrae 33. Four ribs attached to vertebrae 7 to 10.
Pectoral-fin rays seven (i,6); fin margin reaching pelvicfin origin when adpressed. Pelvic-fin rays six (i,5). Dorsal-fin rays eight (i,7); first ray shorter than head length; its origin located dorsal of pelvic-fin base. Anal-fin rays six (i,5). Caudal-fin rays 12 (i,10,i); its distal margin slightly concave; dorsal principal rays longer than ventral rays; in some specimens dorsal unbranched ray slightly elongated, extending distally less than one-third of orbital diameter and not prolonged as filament.
Color in alcohol. Ground color of dorsal surface of adults dark brown; specimens smaller than 85.5 mm SL light brown with small dark spots or vermiculated lines, mainly on snout. Dorsum of head with inconspicuous and scattered dark brown blotches, sometimes forming two weak longitudinal stripes extending from snout tip to anterior region of orbit, passing medially through nares. Longitudinal stripes usually join with greater number of chromatophores laterally and between eyes forming darker region on top of head. Five (4) or six (16*) dark brown transverse bars across body; first on dorsal-fin origin, second on distal margin of dorsal-fin rays, third immediately after vertical line passing through tip of anal-fin rays, fourth and fifth on caudal peduncle, and sixth at end of caudal peduncle, very close to base of caudal fin. Fourth and fifth transverse bars sometimes slightly wider. Fourth and fifth or fifth and sixth bars sometimes join into wider bar (forming five bars instead of six). Some pores of laterosensory system with dark chromatophores, more evident at anterior half of side of body. Color of sensory pores of head indistinct from background. Ventral surface of body pale yellowish. Some specimens with scattered chromatophores on median region of ventral surface of caudal peduncle, at anal-fin origin, and at base of pectoral and pelvic fins.
Fins yellowish with interradial skin hyaline and small darkbrown blotches on its rays. Pectoral fin with interradial skin dark brown, in most specimens its blotches usually expand from rays to interradial skin, mainly on branched portion. Pelvic fin hyaline in most of proximal and medial regions, becoming darker at its branched portion. Dorsal fin with blotches from distal region of first two to four fin rays expanded and joined on interradial skin, forming singular vertical elongated blotch with wider upper region.Anal fin hyaline, with blotches on its distal region. Blotches from distal margin of caudal fin very expanded and joined over interradial skin, forming one or two wide bars occupying most of area. Caudal fin with darkish base.
Rineloricaria maacki View in CoL , new species
Figs. 3-5 View Fig View Fig View Fig
Holotype. MNRJ 31158 View Materials , male, 108.4 mm SL, Brazil, Paraná, União da Vitória, rio Iguaçu , 51°10’31”W 26°10’16”S, 25 May 2003, G. Otto. GoogleMaps
Paratypes. MHNCI 11457 View Materials , 2 ( 1 male, 1 female), 113.0-128.0 mm SL, collected with the holotype GoogleMaps . MHNCI 11455 View Materials , 1 ( female), 102.8 mm SL, Brazil, Paraná, Lapa, areal Água Azul , rio Iguaçu , 50°11’34”W 25°47’34”S, 22 Feb 2001, L. F. Duboc, L. F. S. Ingenito & F. Wegbecher GoogleMaps . MHNCI 11682 View Materials , 1 ( female), 79.7 mm SL, Brazil, Paraná, Lapa, areal Água Azul , rio Iguaçu , 50°11’34”W 25°47’34”S, 10 Nov 2000, L. F. Duboc & L. F. S. Ingenito. GoogleMaps NUP 2540 , 3 ( 1 male, 1 female, 1 c&s female), 110.8-133.5 mm SL, Brazil, Paraná, Cruz Machado / Bituruna , Foz do Areia rerservoir, rio Iguaçu , approx. 51°37’W 26°00’S, 26 Oct 1998, NUPELIA. GoogleMaps NUP 3059 , 1 ( male), 136.2 mm SL, Brazil, Paraná, Cruz Machado / Bituruna , Foz do Areia reservoir, rio Iguaçu , approx. 51°37’W 26°00’S, 6 Oct 1999, COPEL GoogleMaps .
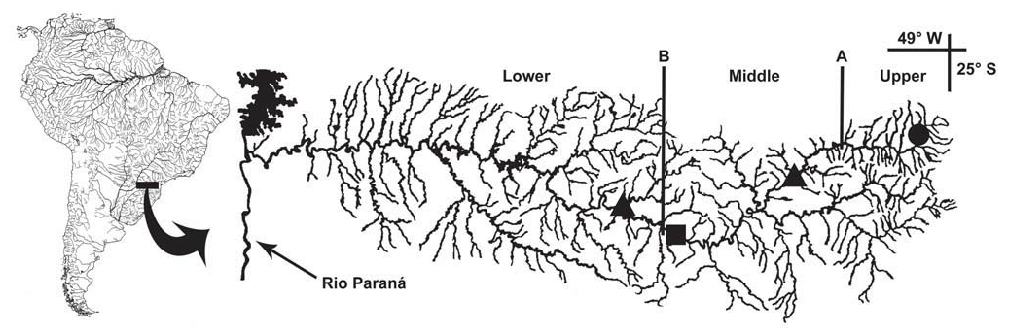
Distribution and habitat. Rineloricaria langei is known from the rio Iraí (a headwater river of upper rio Iguaçu drainage), at the metropolitan region of Curitiba, Paraná State ( Fig. 6 View Fig ). This species inhabits small streams with light to moderate streamflow, over a substrate of sand, some rocks and vegetal debris.
Etymology. The specific name langei is given in honor of Professor Rudolf Bruno Lange, one of the first curators of the zoological collections of Museu de História Natural Capão da Imbuia (MHNCI) during the forties of the Twentieth Century, in which the ichthyological collection is included.
Diagnosis. Rineloricaria maacki is easily distinguished from all Rineloricaria species except from R. latirostris , R. microlepidogaster , R. misionera , R. baliola , R. tropeira , and R. anhaguapitan by having a naked pectoral girdle and the abdomen covered by plates ( vs. pectoral girdle and abdomen completely covered by plates). In addition, the complete covering of plates of the abdominal region of Rineloricaria maacki distinguish it from R. setepovos (that has the abdomen naked); from R. aequalicuspis , R. maquinensis , and R. malabarbai (that have the abdomen variably covered, sometimes naked); and from R. reisi (that has the abdomen almost naked with few irregular platelets). Rineloricaria maacki can be distinguished from R. latirostris by the presence of five dorsal transverse bands ( vs. six bands) and by having the dorsal unbranched caudal-fin ray not prolonged ( vs. dorsal unbranched caudal-fin ray produced as a long filament); from R. misionera , it is distinguished by having the pectoral girdle completely naked ( vs. pectoral girdle with, a few small, well-defined lateral plates). It can be distinguished from both R. anhaguapitan and R. baliola by its snout having a naked area not reaching to anterior most pore of infraorbital ramus of sensory canal species ( vs. snout with a naked area reaching to anterior most pore of infraorbital ramus of sensory canal) and by having all fins, except the dorsal fin, without a distal dark band ( vs. wide dark band covering the distal onehalf of all fins in R. baliola and the same color pattern on dorsal, anal and caudal fins in R. anhaguapitan ); and from R. microlepidogaster , by having abdominal plates between lateral abdominal bones arranged in four ( vs. five to six) series and larger than abdominal plates of this species. Furthermore, R. maacki can be distinguished from R. microlepidogaster by having the pectoral fin usually not reaching the pelvic-fin origin ( vs. pectoral fin surpassing pelvic-fin origin).
Remarks. The paratypes MHNCI 8937 and MNRJ 31157 exhibit an artificial green color, easily seen on ventral surface of the specimens. That unnatural coloration was caused by an artifact during process of fixation and/or preservation in alcohol.
Description. Morphometric data in Table 2 View Table 2 . Head and body deeply depressed. Body depth greatest at dorsal-fin origin. Head triangular in dorsal view. Dorsal profile slightly convex from the tip of the snout to dorsal-fin origin, thereafter gradually depressed up to caudal-fin base. Ventral profile of body nearly straight from snout tip to anal-fin origin, then becoming more depressed towards caudal-fin base. Caudal peduncle depressed forming lateral keels with 11 (3*), 12 (4) or 13 (2) coalesced plates from 28 (1), 29 (4*) or 30 (4) total lateral plates in middle series. Five lateral series of plates, mid-dorsal series present. Scapular bridge completely naked. Abdomen completely covered by plates. Four series of abdominal plates irregularly distributed. Pre-anal plate present and anteriorly rounded by three polygonal plates. Lateral abdominal plates four (3*), five (3) or six (3). Dorsal-fin base with five plate rows.Anal-fin base with two (2) or three (7*) plate rows. Two (2) or three (7*) rows of plates between urogenital pore and anal fin. Plate counts on opposite sides of body usually different, except at dorsal- and anal-fin bases.
Top of head and parieto-supraocciptal smooth. Ridges of parieto-supraocciptal slightly divergent posteriorly. Plates of first three mid-dorsal series without evident ridges, almost smooth. Upper edge of orbit low. Postorbital notch with variable deep, short and narrow, not surpassing one third of the orbital diameter.
Head and body covered by very small odontodes, making fish somewhat smooth. Mature males with small hypertrophied odontodes distributes only on lateral margins of the head and weakly on dorsum of pectoral fin ( Fig. 5 View Fig ). Snout tip with wide naked area, not reaching last pore of infra-orbital ramus of sensory canal. Snout tip ventraly naked, without odontodes between it and upper lip. Lips well developed and covered by papillae; only one irregular papillae row at anterior most area of the upper lip. Two rows of inconspicuous papillae separating upper and lower lips. Maxillary barbel thin and shorter than orbital diameter. Notch present in lower lip. Teeth bicuspid with lateral cusp smallest than medial. Premaxilla with five (2*), six (1), seven (3) or eight (3) teeth.Dentary with five (2), six (1), seven (1) or eight (5*) teeth. Total number of vertebrae 32. Five ribs attached to vertebrae 7 to 11.
Pectoral-fins rays seven (i,6); fin margin sometimes reaching pelvic-fin origin when adpressed. Pelvic-fin rays six (i,5). Dorsal-fin rays eight (i,7); first ray shorter than head lenght; its origin located dorsal of pelvic-fins base. Anal-fin rays six (i,5). Caudal-fin rays 12 (i,10,i); its distal margin slightly concave; dorsal principal rays longer than ventral rays; in two specimens dorsal unbranched ray slightly elongated, extending distally less than one-third of orbital diameter and not prolonged as filament.
Color in alcohol. Ground color of dorsal surface light brown with darker small spots or vermiculated lines. Parietosupraocciptal dark brown. Five dark brown transverse bars across body; first on dorsal-fin origin, second on distal margin of dorsal-fin rays, third near vertical line surpassing through tip of anal-fin rays, fourth and fifth over caudal peduncle. Pores of laterosensory system without evident dark chromatophores. Ventral surface of the body pale or yellowish.
Fins yellowish with interradial skin hyaline and small darkbrown blotches on its rays. Blotches of distal margin of caudal fin very expanded and jointed over interradial skin, forming one bar occupying only its distal third. Caudal fin with darkish base.
Distribution and habitat. Rineloricaria maacki is known from middle and lower rio Iguaçu, a tributary of the rio Paraná basin ( Fig. 6 View Fig ). This species inhabits the main channel and tributaries of rio Iguaçu, which has light brown muddy water with medium to fast flow over a sandy bottom. The habitat has little or no marginal vegetation, mainly formed by grass or bushes.
Etymology. The specific name maacki is given in honor of Reinhard Maack, a geologist who made some of the most important contributions to the knowledge of the geology and physiography of the rio Iguaçu basin and Paraná State.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |