
ASTERIIDAE (Fisher, 1940)
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2010.00688.x |
|
persistent identifier |
https://treatment.plazi.org/id/03E687F8-FFAE-FFA8-3CFE-21A7FE8B5B8B |
|
treatment provided by |
Valdenar |
|
scientific name |
ASTERIIDAE |
| status |
|
THE ASTERIIDAE View in CoL : ANTARCTIC AND
PANTROPICAL CLADES
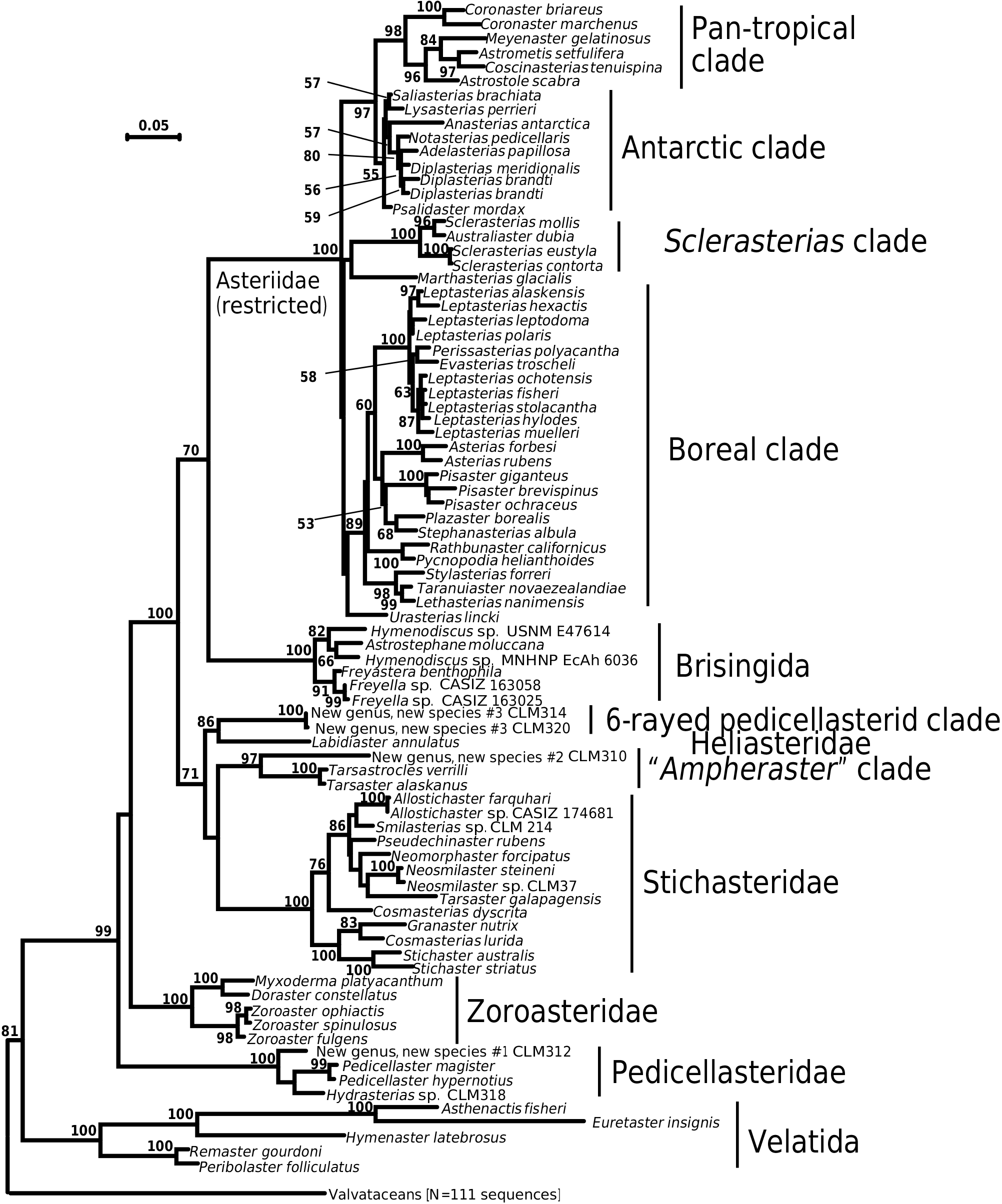
The pantropical asteriid clade was supported as the sister taxon to the Antarctic asteriid clade in Figures 2 View Figure 2 and 3 View Figure 3 , with 94% bootstrap support on the two-gene tree and 97% bootstrap support on the three-gene tree. There is a similar pattern observed in between the Antarctic urchin Sterechinus and its tropical South American sister taxa, Loxechinus and Pseudechinus ( Lee et al., 2004) . This is consistent with a relationship between asteriid taxa from the Antarctic/sub-Antarctic and adjacent tropical regions. Several non-asteriid Forcipulatacea show relationships between tropical and Antarctic/sub-Antarctic taxa. For example, the tropical shallow-water Heliaster is supported as the sister taxon to the Antarctic/ sub-Antarctic Labidiaster . The Antarctic brisingidan Odinella is supported as sister taxon to a clade containing basal brisingidans Brisingaster and Novodinia . Although Novodinia is widely distributed, Brisingaster is known only from the Indian Ocean and New Caledonia ( Mah, 1999). A more complete species-level data set, for both the Antarctic and the pantropical clade, will be needed to discover the closest sister taxon to the Antarctic fauna.
The pantropical Asteriidae is well-supported as the sister group to the diverse Antarctic Asteriidae . The pantropical Asteriidae is composed of Coronaster , members of which occur mostly in the deep sea, as the sister clade to four shallow-water tropical genera: Astrostole , Astrometis , Coscinasterias , and Meyenaster . Astrostole , Astrometis , and Meyenaster are known only from the shallow-water settings from the Eastern Tropical Pacific, whereas Coronaster and Coscinasterias occur widely in the Atlantic and the Indo-Pacific. Coronaster occurs primarily in the deep sea ( > 200 m), although some species (e.g. Coronaster marchenus ) do occur in shallow waters ( < 10 m).
Nearly every known genus of nominal Antarctic Asteriidae was supported as part of a single lineage, occurring only at high latitudes in the Southern Hemisphere. Relative to more stemward Antarctic/ sub-Antarctic forcipulataceans, such as Granaster , Neosmilaster , or Labidiaster , the Antarctic Asteriidae clade evolved more recently and as a single event. Our trees did not support Diplasterias as monophyletic, but Diplasterias forms a species complex in the Southern Ocean region similar to the one formed by Leptasterias in the boreal north. This suggests that further taxonomic sampling and additional genes with longer reads are likely to contribute to a better understanding of Diplasterias , similar to the prior studies of Leptasterias discussed above.
The diversification of the Antarctic Asteriidae may be associated with the isolation of the Southern Ocean fauna and the formation of the Antarctic Counter Current at the end of the Eocene/early Oligocene (25 Ma), as outlined by Clarke & Crame (1992) and Aronson et al. (2009).
Distribution data for several of the genera included in the Antarctic asteriid clade ( Fisher, 1940; Bernasconi, 1973, C. Mah, unpubl. data) show that Anasterias , Diplasterias , Lysasterias , and Psalidaster occur well into the South Atlantic as well as the Southern Ocean, with Adelasterias , Notasterias , and Saliasterias apparently limited to the Southern Ocean. We suggest that the ancestors to the Antarctic asteriid lineage occupied a much broader distribution from the Southern Ocean and the adjacent sub- Antarctic region, including the South Atlantic and South Pacific, prior to the formation of the Antarctic Circumpolar Current (ACC). The ACC would have isolated the Southern Ocean fauna from the adjoining population, leaving the adjoining population ‘outside’ the ACC boundary in the South Atlantic/sub- Antarctic region. Several accounts have documented other invertebrate species that have a similar distribution, but which demonstrate either limited or no gene flow across the ACC (e.g. Hunter & Halanych, 2008; Thornhill et al., 2008). This suggests that nominal Antarctic asteriid species, with identical morphology on either side of the ACC, could be genetically distinct, cryptic species complexes.
The presence of Southern Ocean and sub-Antarctic populations separated by the ACC would also be consistent with widespread circumpolar distribution of so many of the Antarctic asteriids, which lack planktonic larvae. The sub-Antarctic populations would conceivably be ancestral relative to those in the Southern Ocean. This notion would also be consistent with the seeming plesiomorphic morphology shared by Psalidaster , Saliasterias , and several multi-armed and widespread members of the pantropical Asteriidae , such as Coronaster and Astrostole . However, further comprehensive biogeographic analyses of the clade members would be necessary to test this hypothesis.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.