
Herpetocetus (Boessenecker, 2011)
|
publication ID |
https://doi.org/10.5252/g2013n4a5 |
|
DOI |
https://doi.org/10.5281/zenodo.4818791 |
|
persistent identifier |
https://treatment.plazi.org/id/03E7DD69-FFE1-8665-241C-7835FCDA38B3 |
|
treatment provided by |
Felipe |
|
scientific name |
Herpetocetus |
| status |
|
Herpetocetus sp.
REFERRED MATERIAL. — UCMP 219479, a partial juvenile right squamosal, UCMP 219114, a right petrosal with attached compound posterior process, UCMP 219115, a left petrosal lacking posterior process, UCMP 219116, isolated posterior process of a left petrotympanic, UCMP 219117, a partial right tympanic bulla, and UCMP 219077, a partial right mandible, collected by R.W. Boessenecker from UCMP localities V99836 View Materials , V99840 View Materials , and V99859 View Materials .
STRATIGRAPHIC OCCURRENCE. — Middle and upper parts of the San Gregorio section of the Purisima Formation, Early to Late Pliocene ( c. 5-2.5 Ma; Zanclean-Gelasian equivalent; Fig. 2 View FIG ).
DESCRIPTION
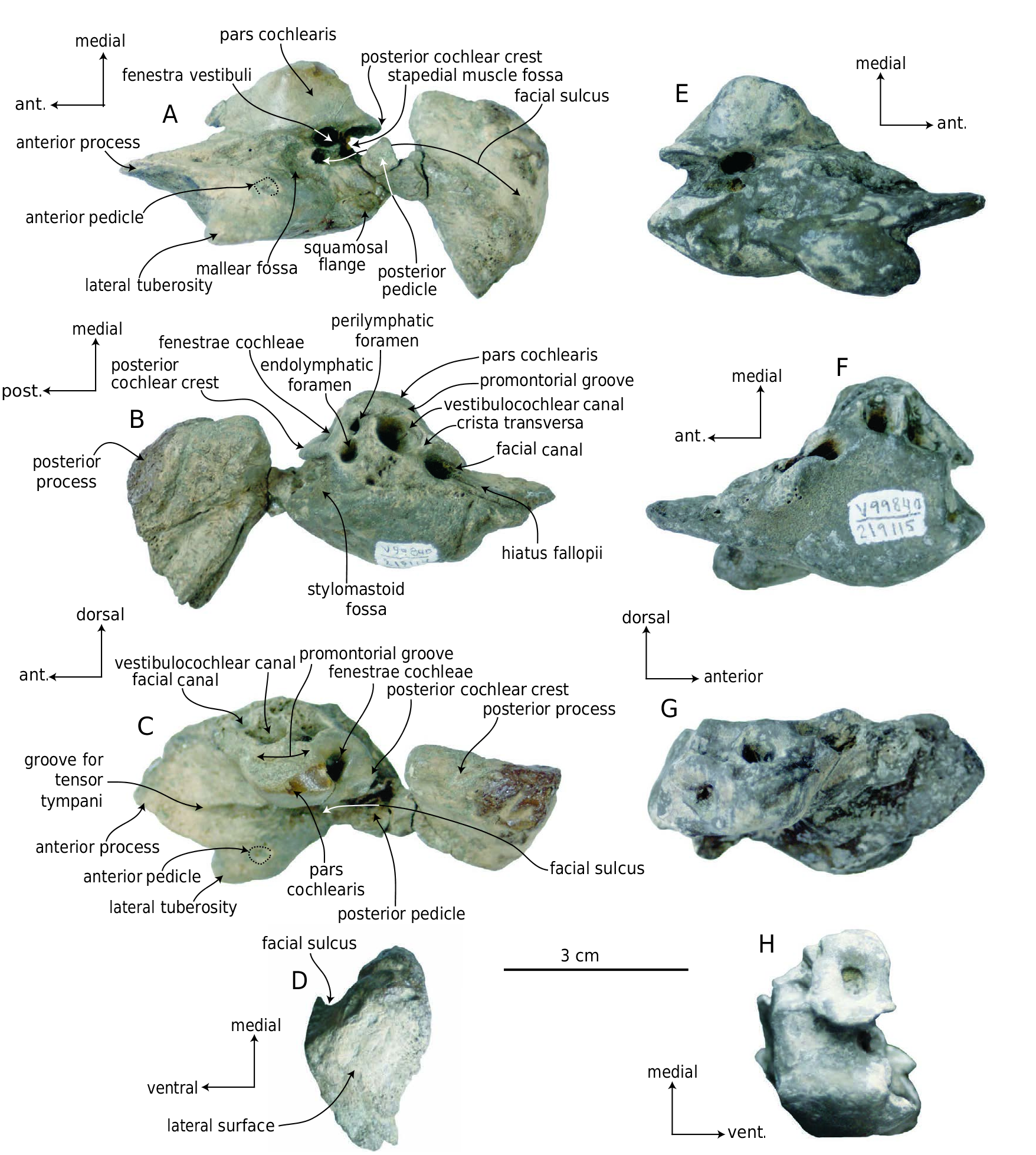
Ŋe squamosal (UCMP 219479) represents a juvenile or neonate based on its small size and lack of fused sutures with adjoining bones (e.g., alisphenoid, parietal, exoccipital). Dorsally, the squamosal is broadly convex where it forms the dorsolateral roof of the braincase and exhibits a longitudinal and elongate fossa along the lateral margin ( Fig. 27 View FIG ). Lateral to the fossa, the supramastoid crest is elongate, sharp, linear, and anteriorly oriented.Two ascending plates of the squamosal extend medial to the supramastoid crest; the squamosal plate extends dorsomedially from the posterior portion of the squamosal, while the falciform process runs anteromedially to form the anterior margin of the braincase ( Fig. 27B View FIG ). Ŋere is a large division between the squamosal plate and falciform process; in more complete crania of Herpetocetus , this wedge-shaped division accommodates the postparietal foramen ( Whitmore & Barnes 2008), the parietal (dorsally) and hypertrophied alisphenoid (anterodorsally). Ŋe zygomatic and postglenoid processes of UCMP 219479 are lost. Only a fragment of the medial edge of the zygomatic is present, but it shows that the process was anteriorly oriented. A knob-like process occurs along the supramastoid crest above the external acoustic meatus. Ŋe sternomastoid fossa is small, concave, triangular, and positioned posterior to the external acoustic meatus and immediately below the supramastoid crest ( Fig. 27D View FIG ). Ŋe lateral surface of the squamosal is otherwise flat. Ventrally, the external acoustic meatus is formed as a transversely oriented concavity posterior to the postglenoid process. Posteriorly, another semi-cylindrical concave fossa is present, oriented posterolaterally, and would have received the posterior process of the petrotympanic. Ŋe medial margin of the squamosal is concave where it forms the lateral wall of the petrosal fossa ( Fig. 27A View FIG ). A corrugated articular surface for the alisphenoid is present dorsomedially.
Ŋe isolated petrosals (UCMP 219114, 219115) are small (pars cochlearis length 29-33 mm; Table 7 View TABLE ) and roughly lenticular in ventral view ( Fig. 28 View FIG ). Ŋe pars cochlearis is small and medially elevated ( Fig. 28A, E View FIG ); the tensor tympani muscle origin is marked by deep groove on the medial surface of the anterior process ( Fig. 28C, F View FIG ). Ŋe anterior process is transversely compressed and bladelike, and appears rectangular in medial aspect.Dorsally adjacent to the groove for the tensor tympani muscle, a longitudinal ridge is present on the medial surface of the anterior process ( Fig. 28C, G View FIG ). Ŋe posterior cochlear crest has a strongly convex posterior margin in medial aspect; in ventral aspect it is well developed, and strongly overhangs both the stapedial muscle fossa and the stylomastoid fossa ( Fig. 28C, G, H View FIG ). Ŋe dorsal side of UCMP 219115 is convex and lacks a suprameatal fossa, while in UCMP 219114, it is nearly planar with a shallow suprameatal fossa. Ŋe canal for the vestibulocochlear nerve is large and circular, and is separated from the facial canal by a thick crista transversa ( Fig. 28B, F View FIG ). In dorsal aspect, the slit-like hiatus fallopii is confluent with the anterior margin of the facial canal, giving the facial canal a teardrop shape. Relative to the endolymphatic foramen, the perilymphatic foramen is much smaller ( Fig. 28B View FIG ); both are oriented slightly anterodorsally. At the position of the anterior part of the pars cochlearis, the lateral tuberosity is present along the ventral margin of the anterior process; it is ventrolaterally projecting as a flattened triangular flange in UCMP 219114 ( Fig. 28A View FIG ) and a large knob in UCMP 219115 ( Fig. 28E View FIG ). In UCMP 219115, a short groove (that widens and deepens anteriorly) is present along the anterior half of the lateral tuberosity, separating the lateral tuberosity and anterior process. Medial to the lateral tuberosity the broken anterior pedicle occurs as a small, anteroposteriorly short and bladelike crest. Between the anterior pedicle and the pars cochlearis is a relatively large, shallow mallear fossa. Ŋe fenestra vestibuli is oval and positioned posteromedial to the mallear fossa; the facial canal opens lateral to the fenestra vestibuli, and opens into the facial sulcus. Ŋe facial sulcus and fenestra vestibuli are separated by a thin crest ( Fig. 28A View FIG ). Posterior to the lateral tuberosity, a small convex ridge-like squamosal flange ( sensu Geisler & Luo 1996) is ventrally prominent. Ŋe lateral surface of the petrosal is broadly convex in both specimens ( Fig. 28A, B, E, F View FIG ). Medially adjacent to the squamosal flange, the posterior pedicle is preserved in both specimens as a transversely thin crest.
Ŋe posterior process (UCMP 219115, 219116) is short ( Table 8 View TABLE ) and plug-shaped ( Figs 28 View FIG A-D; 29D- F); in UCMP 219115, there is a thin, constricted neck. Ŋe lateral surface of the posterior process is flat, triangular, and slightly rugose. Ŋe dorsolateral apex is anteroposteriorly tapered ( Figs 28D View FIG ; 29F View FIG ). Ŋe short, deep facial sulcus occurs on the ventral side, and is bounded anterolaterally by a large, high, bladelike ridge ( Figs 28D View FIG ; 29F View FIG ). Posteromedial to the facial sulcus is an anteroposteriorly thick, transversely oriented ridge, which is anteromedially convex and spherical. Medially adjacent to the neck of the posterior process is a small but well defined circular pit.
The partial right tympanic bulla (UCMP 219117) is abraded and missing most of the outer lip ( Fig. 29 View FIG A-C; Table 5 View TABLE ). Ŋe involucrum is relatively thick with a well developed dorsal posterior prominence ( Fig. 29B, C View FIG ). In ventral aspect, the bulla is posteriorly bilobate: a fossa-like median furrow separates the medial and lateral lobes. Ŋe involucral and main ridges are posteriorly divergent and widely separated ( Fig. 29C View FIG ). In medial aspect, the anterior portion of the involucrum is constricted, and the medial surface of the involucrum bears fine transverse striations.
Ŋe anterior half of the mandible (UCMP 219077) is missing, but the posterior portion is well-preserved ( Fig. 30 View FIG ; Table 6 View TABLE ) and similar to UCMP 219079 with a few exceptions ( Table 6 View TABLE ). Laterally adjacent to the dorsal crest, three mental foramina are preserved and open into anteriorly extending sulci ( Fig. 30A View FIG ). Two small gingival foramina occur on the medial surface, medially adjacent to the dorsal crest anterior to the coronoid process, also anteriorly opening but with shorter sulci than the gingival foramina. A large, deep, and anteroposteriorly elongate fossa on the lateral surface of the coronoid process extends anteriorly as a narrow concavity for the temporalis insertion below the anterior crest of the coronoid process. Ŋe mandibular condyle is laterally bilobate, with a slight fossa occurring on the lateral side dividing the condyle into dorsal and ventral lobes. A deep fossa occurs on the ventral
A coronoid process elevation inward mandibular condyle
side of the condyle, forming a sharp ventromedially curving crest ( Fig. 30D View FIG ). Ŋe angular process extends far posterior to the condyle and dorsally exhibits a longitudinal groove.
COMPARISONS
Ŋese specimens can be referred to Herpetocetus owing to several characteristics.Ŋe squamosal (UCMP 219479) matches those of Herpetocetus as described by Whitmore & Barnes (2008) in possessing a large fossa for the posterior process of the petrotympanic, an elongate anteroposteriorly oriented trough-like fossa on the squamosal medial to the supramastoid crest, and a large wedge-shaped dorsomedial margin to accommodate the alisphenoid and postparietal foramen. Ŋe squamosal differs from Herpetocetus bramblei in exhibiting a small sternomastoid fossa that does not extend anterior to the level of the external acoustic meatus. Ŋe petrosals bear a laterally compressed, bladelike anterior process (which characterizes Herpetocetus and Nannocetus Kellogg, 1929 ; Geisler & Luo 1996; Whitmore & Barnes 2008), a large lateral projection ( sensu Geisler& Luo 1996; = lateral tuberosity, Mead & Fordyce 2009) of the anterior process (unique to Herpetocetus ; Geisler & Luo 1996; Whitmore & Barnes 2008), and a short plug-shaped posterior process (a potential synapomorphy of herpetocetines; Bouetel & Muizon 2006; Steeman 2007; Whitmore & Barnes 2008). Ŋe squamosal (UCMP 219479) was placed in articulation with UCMP 219114 ( Fig. 27C View FIG ), a petrosal with attached posterior process from a separate individual (see description above); the tight articulation of these two elements strongly supports the identification of these as the same taxon.
Ŋe tympanic bulla (UCMP 219117) is referred to Herpetocetus sp. because of its small size, distinct dorsal posterior prominence, median groove, distinct transverse creases on the involucrum, and close phenetic similarity to previously published tympanic bullae of Herpetocetus ( Whitmore & Barnes 2008) . A newly referred (and nearly complete) skull of H. bramblei (UCMP 219111) introduced by Boessenecker & Geisler (2008) includes both petrosals and a tympanic bulla, the latter of which is not preserved in the holotype. Ŋe isolated tympanic bulla (UCMP 219117) differs from the well-preserved tympanic bulla of UCMP 219111 in its smaller size, less distinct transverse creases, much lower dorsal posterior prominence, a shallower furrow on the medial surface between the dorsal prominence and the posterior lobe, and lacking a sharp spur on the ventral tip of the dorsal posterior prominence. UCMP 219117 differs from Herpetocetus transatlanticus Whitmore & Barnes, 2008 , the referred specimen of H. “ sendaicus ”, and the holotype bulla of H. sendaicus , in having a transversely narrower involucrum with a smaller and more defined dorsal posterior prominence. It also differs from Piscobalaena and Nannocetus in exhibiting a more strongly developed dorsal posterior prominence and having a more constricted anterior portion of the involucrum.
Ŋe mandible (UCMP 219077) exhibits characteristic features of Herpetocetus , including an elongate and laterally hooked coronoid process, a posteromedial sloping mandibular condyle, and a posteriorly projecting and shelf-like angular process (see above remarks under H. bramblei ; Boessenecker 2011b; Whitmore & Barnes 2008). Ŋis specimen is unique among herpetocetines (and possibly among mysticetes) in having a fossa on the lateral surface of the condyle, giving it a bilobate morphology. It is further distinct from Piscobalaena , H. “ sendaicus ”, H. bramblei , and indeterminate herpetocetine mandibles ( Boessenecker 2011b) in having a fossa for the temporalis insertion that extends anteriorly beyond the coronoid process; this feature is slightly developed in H. scaldiensis . UCMP 219117 is roughly the same size as the mandible of H. “ sendaicus ”, and is much smaller than UCMP 219076 described above. As mentioned above, the mandibular morphology of herpetocetines may not be diagnostic at the genus level ( Boessenecker 2011b), but some subtle mandibular features may be taxonomically useful among species of Herpetocetus .
REMARKS
Ŋese specimens appear to represent an undescribed species of Herpetocetus morphologically distinct from and geochronologically younger than H. bramblei ; additional material of this taxon is recorded from the Santa Cruz section of the Purisima Formation (see below). Ŋe squamosal, petrosals, and mandible exhibit several differences with H. bramblei (see above). Ŋese specimens are from higher up in the San Gregorio section than the mandible of H. bramblei (UCMP 219079), and occur in younger strata with occasional Patinopecten healelyi shells, the latter indicating a Pliocene age (Powell et al. 2007). Ŋe mandible (UCMP 219077) was collected from between two tephra layers dated at 2.5 and 3.35 Ma, indicating an Early to Late Pliocene age (Piacenzian- Gelasian equivalent) for this specimen. All other specimens were collected below the 3.35Ma tephra, and above the aforementioned 6.4-5.6 Ma (Late Miocene) diatom flora; they are likely Early-Late Pliocene (Zanclean-Piacenzian equivalent) in age due to their association with Pliocene mollusks and Carcharodon carcharias teeth ( Boessenecker 2011a). Ŋese specimens are all younger than the 6.79- 5.33 Ma age for the type locality of H. bramblei . Likewise, these specimens differ from H. bramblei . Ŋe squamosal (UCMP 219479) exhibits a small sternomastoid fossa that does not extend anteriorly past the external acoustic meatus ( Fig. 27D View FIG ); in H.bramblei , the sternomastoid fossa is much larger. A similarly small sternomastoid fossa occurs in a Herpetocetus sp. braincase (UCMP 219121) from the Lower Pliocene Purisima Formation near Santa Cruz (5.33-4.5 Ma; Zanclean equivalent). Ŋe petrosals (UCMP 219114 and 219115) are relatively similar to the holotype of H. bramblei , but both differ from it in having a posteriorly elongate and transversely thin posterior cochlear crest, and a thick crista transversa; the crista transversa in the H. bramblei holotype is thin, and the posterior cochlear crest is short. Ŋe H. bramblei holotype petrosal further differs in having elongate prongs of bone on the dorsal surface, and having a circular facial canal which lacks an anterior fissure (as opposed to the teardrop shaped facial canal described above in UCMP 219114 and 219115). UCMP 219115 also differs from H.bramblei (and all other herpetocetines) in exhibiting a bulbous lateral projection of the anterior process ( Fig. 28E View FIG ).
Ŋese specimens are distinct from H. bramblei , and indicate the presence of a second, younger undescribed species within the Purisima Formation. Additional material from the lower Pliocene section of the Purisima Formation (Zanclean-Piacenzian equivalent) near Santa Cruz includes a skull with earbones (UCMP 219121) and several isolated petrosals (UCMP 219118, 219119) that share this same morphology. As this more diagnostic material is from a locality separate from material described herein, it is beyond the scope of this study to describe this new taxon, and will be considered in a separate study.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.