
Probolodus sazimai, Santos & Castro, 2014
|
publication ID |
https://doi.org/10.1590/1982-0224-20130232 |
|
DOI |
https://doi.org/10.5281/zenodo.5137673 |
|
persistent identifier |
https://treatment.plazi.org/id/03E83A01-FFB7-FFDB-FE86-7B92FDC9FBCB |
|
treatment provided by |
Carolina |
|
scientific name |
Probolodus sazimai |
| status |
sp. nov. |
Probolodus sazimai View in CoL , new species
Figs. 7-9 View Fig View Fig View Fig
Probolodus heterostomus, Eigenmann, 1911: 164 View in CoL [in part]. - Eigenmann, 1915: 20-22 [in part]. - Sazima, 1977: 510 [in part] (distribution). - Bizerril, 1994: 58, 65 [in part] (distribution and identification key). - Lima et al., 2003: 154 [in part] (type-material, type locality and distribution). - Oyakawa et al., 2006: 156 [in part] (distribution). - Buckup et al., 2007: 61 [in part] ( type locality and distribution). - Menezes et al., 2007: 105 [in part] ( type locality; distribution; systematic and ecology).
Holotype. MCP 47870, 56.7 mm SL, Brazil, Minas Gerais, município de Ipatinga, rio Taquaruçu, tributary of rio Doce basin, 19°27’S 42°28’59"W, 09 Nov 1989, C. Lucena, E. Pereira, J. Pezzi & P. Azevedo. GoogleMaps
Paratypes. All specimens from Brazil. Espírito Santo State : MZUSP 27577 View Materials , 1 View Materials , 46.5 mm SL, rio Itapemirim basin, fazenda Boa Esperança between the towns of Pacotuba and Coutinho , 20°45’S 41°18’59"W GoogleMaps ; MZUSP 79668 View Materials , 1 View Materials , 50.7 mm SL, município de Linhares, rio Doce basin, Juparanã pond, 19°23’28"S 40°04’20"W GoogleMaps . Minas Gerais State: LBP 3433 , 1 , 45.1 mm SL, rio Doce drainage basin, rio Matipó , 20°18’09"S 42°20’04"W GoogleMaps ; MCP 13813, 4 View Materials , 36.8-54.4 mm SL, município de Ipatinga, rio Doce basin, rio Taquaruçu along
road BR-381, 19°27’S 42°28’59"W (1 c&s, 43.5 mm SL), collected with holotype; MNRJ 17120 View Materials , 1 View Materials , 36.7 mm SL, município de Chiador, córrego da Tocaia, left hand tributary of rio Paraíba do Sul, 22°00’10"S, 43°03’28"W GoogleMaps ; MZUSP 41725 View Materials , 3 View Materials , 40.7-51.7 mm SL, município de Ipatinga, rio Doce basin, rio Taquaruçu along road BR-381, 19°39’59"S, 42°26’00"W GoogleMaps .
Diagnosis. Probolodus sazimai differs from its two congeners, P. heterostomus e P. oyakawai (new species) by the following characteristics: six transverse series of scales from insertion of pelvic fin to lateral line ( vs. seven/eight); 12 scales along middorsal line between posterior termination of base of dorsal fin and adipose fin ( vs. 14-17 and 13-16, respectively); presence of two pairs of uroneurals ( vs. absent or just one pair) ( Fig. 8a View Fig ); anterior margin of the hypurals 2 and 3 in contact, not fused ( vs. hypurals 2 and 3 distinctly separate) ( Fig. 8a View Fig ); end piece or stay of the dorsal fin leaning against or fused in the last pterygiophore ( vs. somewhat removed of the last pterygiophore) ( Fig. 8b View Fig ); fourth basibranchial completely cartilaginous ( vs. ossified) ( Fig. 9a View Fig ); dorsal margin of the opercle more elevated than the hyomandibular ( vs. opercle and hyomandibular practically aligned) ( Fig. 9b View Fig ); anteromedially projection of the mesethmoid wide, separating completely the premaxillary ( vs. short projection, not separating completely the premaxillary). It may be further separated from P. heterostomus by presence of 36-43 perforated scales on the lateral line ( vs. 45-56 perforated scales).
Description. Morphometric data presented in Table 1 View Table 1 . Body moderately compressed and deep, less so in individuals of less than 150 mm SL, greatest body depth within region delimited anteriorly by vertical through insertion of pelvic fin and posteriorly by vertical through origin of dorsal fin. Average body depth variable among population samples, but with broad overlap in ranges among examined samples. Dorsal profile of head distinctly convex from margin of upper lip to vertical through posterior nostril, straight to very slightly convex from latter point to tip of supraoccipital spine. Dorsal profile of body moderately convex from tip of supraoccipital spine to origin of dorsal fin, straight and posteroventrallyslanted along base of dorsal fin, straight to slightly convex from posterior terminus of base of dorsal fin to adipose fin, and slightly concave along caudal peduncle. Broad middorsal ridge present along predorsal region of body, with ridge less obvious anteriorly. Dorsal region of body between posterior terminus of dorsal fin and adipose fin transversely rounded overall, but slightly flattened middorsally in some individuals. Ventral profile of head slightly convex anteriorly from margin of lower lip approximately to vertical through anterior nares, then convex from that point to vertical through posterior margin of eye. Ventral profile of body convex to insertion of pelvic fin, nearly straight but slightly posteroventrally-aligned from that point to origin of anal fin, straight to slightly convex and posterodorsally-slanted along base of anal fin, and slightly concave along caudal peduncle. Prepelvic region of body somewhat flattened transverselly.
Head obtusely rounded anteriorly in lateral profile. Mouth terminal. Upper jaw with maxilla distinctly posteroventrallyangled relative to ventral margin of premaxilla and extending ventral of orbit to point somewhat short of, or reaching, vertical through anterior margin of pupil. Nares of each side of head very close together; anterior opening circular, posterior crescent-shaped with distinct flap between nares. Eye relatively large and lacking distinct adipose eyelid. Median fronto-parietal fontanel extending from mesethmoid to supraoccipital spine. Width of fontanel approximately onefifth distance across bony interorbit. Infraorbital series complete with third infraorbital by far largest. All infraorbitals carrying laterosensory canal segments proximate to margin of orbital rim. Supraorbital absent. Branchiostegal rays four. Gill-rakers long and setiform; 10-12+1+7-8 rakers present on first gill-arch.
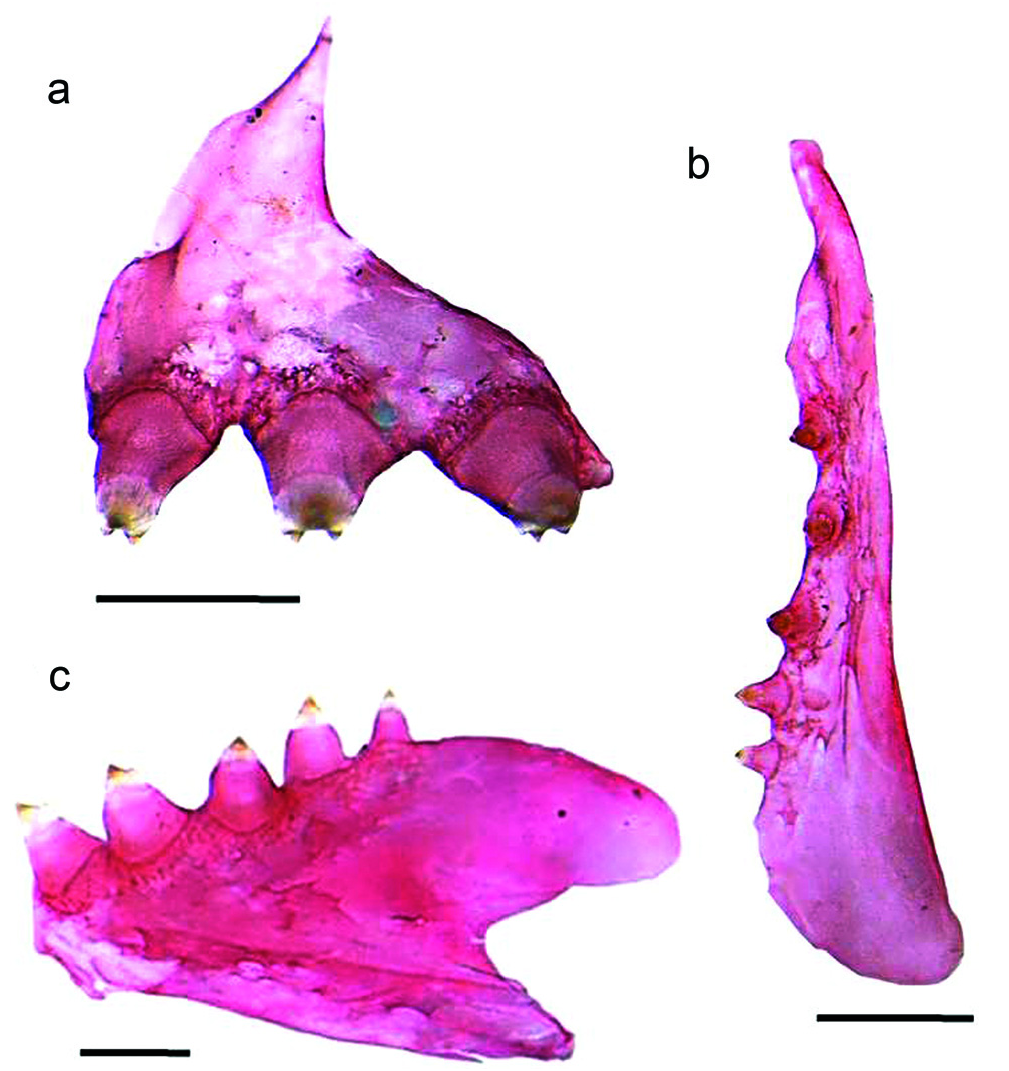
Teeth on premaxillary, maxillary, and dentary in one row. Premaxilla with three (9) or four (2) teeth [three] somewhat directed outward; each tooth with three points in angles of a nearly isosceles triangle; middle point, which is also the anterior one, much larger. Maxilla with three to six (commonly four) teeth [four] with two cusps of similar size, first two or three directed outward. Dentary, mostly, with six large teeth, first three directed outward, fourth, fifth, sometimes the sixty tricuspidate, and one or more smaller ones following it, directed upward; larger teeth of the lower jaw heavy, conical, with a minute cusp on each side; in overall, dentary has five to six (commonly five) tricuspidate teeth [five].
Scales cycloid, relatively large, and firmly implanted. Lateral line decurved anteriorly approximately to vertical through origin of dorsal fin, then slightly posterodorsallyangled to caudal peduncle, and then running horizontally to terminus of scale series. Lateral line completely pored from supracleithrum to base of caudal fin and followed posteriorly by assified tubular extension running along lateral margin of membrane joining middle rays of caudal fin. Lateral-line scales 36 (2), 38 (2), 39 (1), 40 (3), or 43 (2) [42]; scales in transverse series from origin of dorsal fin to lateral line 7 (10) or 8 (1) [7]; scales in transverse series from insertion of pelvic fin to lateral line 6 (11) [6]; scales in transverse series from origin of anal fin to lateral line 6 (1) or 7 (10) [7]; scales along middorsal line between tip of supraoccipital process and origin of dorsal fin 12 (9) or 13 (2) [12]; scales along middorsal line between posterior termination of base of dorsal fin and adipose fin 12 (11) [12]; horizontal scale rows around caudal peduncle 14 (6), 15 (1), or 16 (2) [16]. Vertebrae 33 (n = 4), 34 (n = 4), 35 (n = 2), or 36 (n = 1) [34].
Dorsal-fin rays ii,9 (11) [ii,9]. Anal-fin rays iv (7) or v (4), 22 (1), 23 (2), 24 (4), 25 (1), 26 (2), or 27 (1) [v,25]. Pectoral-fin rays i,10,i (4), i,11 (1), i,11,i (5), or i,12 (1) [i,10,i]. Pelvic-fin rays i,6 (2), i,6,i (1), or i,7 (7) [i,6,i]. Principal caudal-fin rays i,17,i (11) [i,17,i].
Dorsal-fin margin distally rounded to slightly truncate; first unbranched ray approximately 40-50% length of second unbranched ray. Dorsal-fin origin situated along vertical located approximately at middle of SL. Origin of adipose fin located slightly anterior of vertical through posterior terminus of base of anal fin. Profile of adpressed pectoral fin distinctly acute. Tip of pectoral fin extending approximately to vertical through insertion of pelvic fin. Profile of expanded pelvic fin pointed, with first branched ray either longest in fin or subequal to unbranched ray. Insertion of pelvic fin located distinctly anterior to vertical through origin of dorsal fin. Tip of adpressed pelvic fin extending approximately to vertical through insertion of anal fin. Some larger, apparently male, specimens with posteriorly-directed hooks along posterior margins of second through seventh pelvic-fin rays; hooks rarely also present on first ray. Hooks limited to posterior branch of hook-bearing rays. Each hook-bearing segment typically with one hook, although two hooks occasionally present on some segments. Distal margin of anal fin slightly concave in which fourth or fifth unbranched and first and second branched rays longest and subequal or first through third branched rays longest with subsequent branched rays gradually decreasing in length. Some larger, apparently male, individuals with hooks present on various rays gradually of anal fin. Hooks most often arise along posterior margin of posterior branch of second through seventh branched rays. Hooks extending onto unbranched segments of some rays. Caudal fin forked, with distal margins of lobes obtusely pointed.
Color in alcohol. Overall ground color of body in specimens fixed in formalin yellowish brown. Guanine remaining on lateral and ventral portions of head and on ventral and, to a variable degree, lateral surfaces of body. Snout and dorsal portion of head relatively dark. Middorsal and immediately adjoining portions of body relatively dark. Distinct, wedge-shaped, ventrally-attenuated humeral blotch with irregular margins extending from approximately five scales dorsal of lateral line to about two scales ventral of lateral line. Pigmentation of humeral blotch typically most intense in region dorsal of lateral line, with pigmentation of remaining ventral portion of blotch variably lighter, but still quite obvious. Dark midlateral pigmentation more conspicuous on portion of body beginning at vertical through anterior portion of dorsal fin, with stripe widening posteriorly on caudal peduncle into distinct, horizontally-elongate, ovoid blotch. Pigmentation of blotch more intense than that of remainder of stripe. Chromatophores of dorsal portion are concentrated at the base and middle region of the scales, its margins being free of chromatophores. Region below the lateral line decreases the concentration of chromatophores, and, between the anal fin and caudal peduncle, sparse chromatophores are present in greater proportion.
Dorsal, anal, and caudal fins with interradial membranes covered by small dark chromatophores, with chromatophores often more concentrated proximate to margins of fin rays. Dark pigmentation on caudal fin more intense along middle fin-rays in most specimens and forming stripe variably continuous anteriorly with midlateral dark blotch on caudal peduncle. Anal fin with unbranched rays and all branched rays pigmented. Distal half of remainder of fin with dark chromatophores in all individuals, with fields of chromatophores extending nearly to base of fin membranes. Adipose fin lightly colored overall, but often freckled with small dark spots. Pectoral and pelvic fins with small, dark spots located both along fin-ray margins.
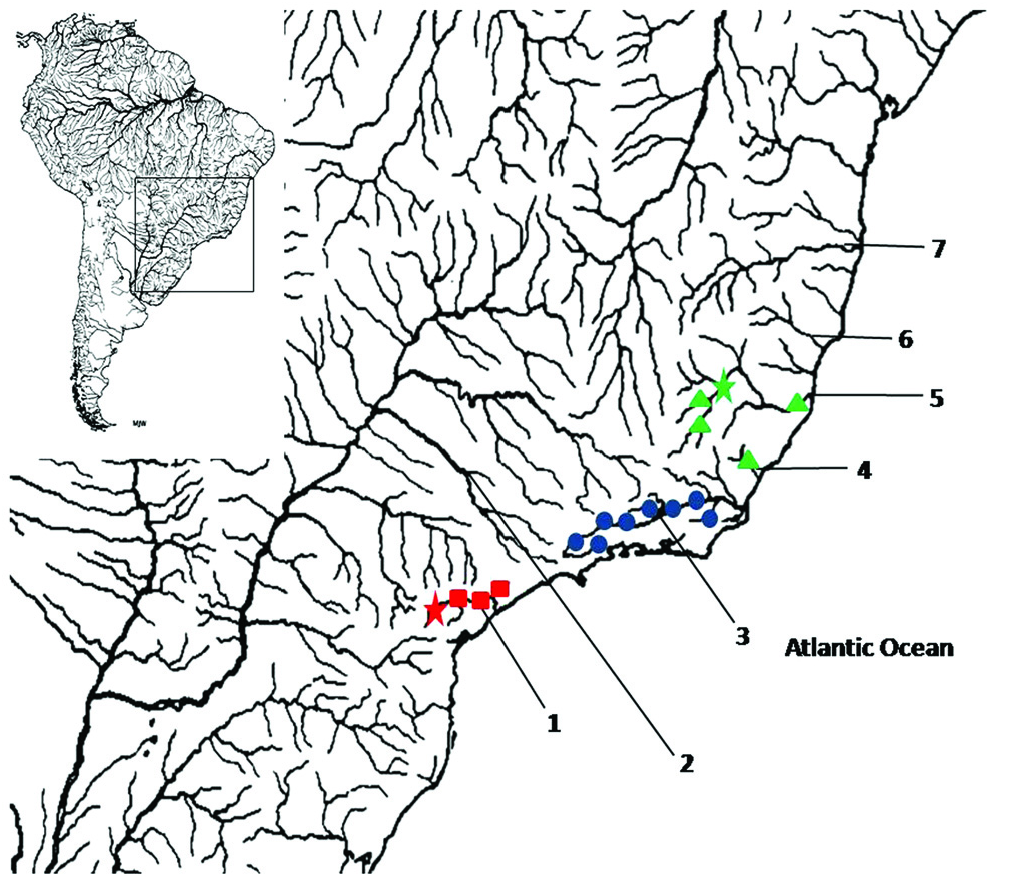
Distribution. Probolodus sazimai is known to occur in the rios Itapemirim and Doce basins ( Espírito Santo and Minas Gerais states, respectively), southeastern Brazil. ( Fig. 2 View Fig ).
Ecological notes. Ctenoid and cycloid fish scales were observed in the stomach contents and between gill rakers of one specimen (MCP 13813, 1, 43.5 mm SL). No other items could be identified, possibly due to their advanced state of decomposition. Leaf fragments, insects and scales were observed in the oral cavity of three specimens preserved in alcohol (LBP 3433, 1, 45.1 mm SL; MCP 13813, 2, 36.8-54.4 mmSL).
Etymology. The species name, sazimai , is in honor of Ivan Sazima from Museu de Zoologia of UNICAMP (ZUEC-PIS), in appreciation of his contributions to our knowledge of the Neotropical lepidophagous caraciforms.
Notes on phylogenetics relationships and biogeography of Probolodus
The monophyly of Probolodus is supported by five exclusive characteristics, and three non-exclusive characteristics. Of all the characteristics shared among species of the genus, there are at least two exclusive characteristics, such as the presence on premaxillary and dentary of tricuspid teeth, with median cusp greatly developed and pointed outward, most probably related to the lepidophagous habit these species ( Fig. 1 View Fig a-c).
The genera of Neotropical characiforms Bryconexodon , Catoprion , Exodon , Roeboexodon , Roeboides , and Serrabrycon also exhibit specialized dentitions favoring the lepidophagous habit, such as pointed by Sazima (1977), Sazima & Uieda (1980), Sazima & Machado (1982) and Sazima (1983). However, specialized teeth directed out of the mouth is a diagnostic characteristic which allows easy recognition of lepidophagous taxa, although, the shape, number and the differential arrangement these conical and mammiliform, or cuspidate teeth on premaxillary, maxillary and dentary in these species are fundamental for understanding of evolution this character in independent lineages of Neotropical lepidophagous characiforms, as demonstrated by some authors ( e.g., Roberts, 1970; Géry, 1980; Sazima, 1983; Vari, 1986; Lucena, 1998; Mirande, 2010; Mattox & Toledo-Piza, 2012). Thus, the specialized dentition of Probolodus , as well as the predatory behavior of plucking and eating scales could have evolved independently in Bryconexodon , Exodon , and Roeboexodon . Among the shared characteristics that support evolutionary relationships among these three taxa, we highlight some related to the structure and arrangement of teeth, mainly: the teeth of these characids are stout with hypertrophied bases, conical, and especially in the dentary, the larger teeth have compressed and cuspidate crowns, being unicupidate or bicuspidate and directed upwards; larger cuspidate teeth arranged with conical smaller teeth, and are recurved posteriorly relative to the body; in the premaxillary there is a outward mammiliform tooth developed and directed forward.
Probolodus has been treated as a monophyletic group of lepidophagous characids evolutionarily related to the Tetragonopterinae ( sensu Roberts, 1970; sensu Géry, 1977; 1980; and sensu Mirande, 2010). Thus, the hypothesis of Sazima (1977), considers the similarity of Probolodus with the Tetragonopterinae ( sensu Géry, 1977) a form of aggressive mimicry, ( i.e., similarity due to mimicry and not to phylogenetic proximity), whereas the Tetragonopterinae ( sensu Géry, 1977), Astyanax fasciatus and coastal sympatric species of the genus Deuterodon are probably the usual prey of the species of Probolodus .
The species of the genus Probolodus are distributed throughout the coastals drainages of eastern Brazil (from Espírito Santo to São Paulo states) which according to Ribeiro (2006) corresponds to an area of great biogeographical significance due to high degree of endemism in its fish fauna ( Fig. 2 View Fig ). Meanwhile, the eastern coastal systems have already been considered as areas of endemism by various authors ( e.g., Eigenmann, 1909; Géry, 1969; Vari, 1988; Weitzman et al., 1988; Menezes, 1996) and recognition of these areas in the coastal drainages of South and Southeast of Brazil is similar or quite congruent with the distribution of the genus Deuterodon by Lucena & Lucena (1992; 2002) and Pereira (2010), Mimagoniates by Menezes & Weitzman (1990), Spintherobolus by Weitzman & Malabarba (1999), Hemipsilichthys by Pereira & Reis (2002), Hollandichthys by Bertaco (2003), Phalloptychus by Lucinda (2005), Phalloceros by Lucinda (2008).
However, the coastal streams have been historically connected with the adjacent upland crystalline shield. Thus, geological events were responsible to the capture of adjacent upland high areas, and consequently the capture of upland streams, such as the formation of the upper rio Paraíba do Sul by capturing streams of the rio Tietê ( cf., Ab’Saber, 1957; Menezes, 1972; Weitzman et al., 1988; Langeani, 1989; Bizerril, 1994; M. Malabarba, 1998; Weitzman & Malabarba, 1999; Bizerril & Lima, 2000; Oliveira & Britski, 2000; Quevedo & Reis, 2002; Oyakawa et al., 2005; Ribeiro et al., 2006; Lucinda, 2008). Therefore, possible effects of tectonics, changing the topography of the terrain and, thus, influencing the capture of coastal streams, especially in the southeastern (Ab’Saber, 2003; Cobbold et al., 2001; Ribeiro, 2006) and fluctuations in sea level in the Pleistocene, allowing the union of several rivers and coastal streams, favoring the isolation of populations and speciation events ( Weitzman et al., 1988; Weitzman & Malabarba, 1999; Ribeiro, 2006), are, probably, related to the divergence of the species of Probolodus .
Comparative material examined. Aphyocharax pusillus, LIRP 4559, 2, 39.6-45.4 mm SL; Astyanax fasciatus, LIRP 132, 3, 71.9- 75.4 mm SL; Brycon pesu, LIRP 5962, 2, 73.3-82.9 mm SL; Bryconexodon juruenae, LIRP 7139, 2, 89.8-98.8 mm SL; Bryconops alburnoides, LIRP 5116, 2, 94.0- 110.7 mm SL; Cheirodon galusdae, LIRP 5958, 3, 38.7-45.0 mm SL; Deuterodon iguape, LIRP 1059 , 2, 83.2-86.5 mm SL; Exodon paradoxus, LIRP 5933, 2, 59.6-62.8 mm SL; Gymnocorymbus thayeri, MZUSP 17468, 2, 44.8-46.6 mm SL; Hemigrammus unilineatus, MZUSP 30322, 2, 30.1-33.2 mm SL; Hollandichthys multifasciatus, LIRP 5744, 2, 70.6-86.5 mm SL; Hyphessobrycon eques, LIRP 1835 , 2, 29.6-30.9 mm SL; Jupiaba anteroides, MZUSP 17929, 3, 63.6-74.4 mm SL; Moenkhausia xinguensis, MZUSP 91690, 2, 59.8-64.6 mm SL; Oligosarcus pintoi, LIRP 300, 2, 75.2-82.06 mm SL; Poptella paraguayensis, LIRP 5940, 2, 35.2-39.8 mm SL; Roeboexodon guyanensis, MZUSP 91949, 2, 83.4-83.8 mm SL; Tetragonopterus argenteus, LIRP 5941, 2, 49.7-56.1 mm SL.



| MCP |
Pontificia Universidade Catolica do Rio Grande do Sul |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Probolodus sazimai
| Santos, Osmar & Castro, Ricardo M. C. 2014 |
Probolodus heterostomus , Eigenmann, 1911: 164
| Buckup & Menezes & Ghazzi & Catalogo 2007: 61 |
| Buckup & Menezes & Ghazzi & Catalogo 2007: 105 |
| Oyakawa 2006: 156 |
| Lima 2003: 154 |
| Bizerril 1994: 58 |
| Sazima 1977: 510 |
| Eigenmann 1915: 20 |
| Eigenmann 1911: 164 |