
Microtechnites, BERG, 1879
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00770.x |
|
persistent identifier |
https://treatment.plazi.org/id/03E8878D-FFDB-FFDB-5EFD-FB63B7BCFA23 |
|
treatment provided by |
Marcus |
|
scientific name |
Microtechnites |
| status |
|
MICROTECHNITES BERG View in CoL View at ENA ( FIGS 4 View Figure 4 , 37–38 View Figure 37 View Figure 38 )
Microtechnites Berg, 1883: 73 View in CoL [gen. nov.; type species Capsus pygmaeus Berg, 1879 View in CoL (junior homonym of Capsus (Deraeocoris) pygmaeus Berg, 1878 View in CoL ) = Halticus spegazzinii Berg, 1883 by monotypy]; Reuter, 1910: 149 (cat.); Carvalho, 1952: 74 (synonymy with Halticus Hahn View in CoL ); Carvalho, 1958: 13 (cat.); Cassis & Gross, 1995: 186 (cat.; synonymy); Schuh, 1995: 53 (cat.; synonymy). Reinstated status.
Cafayatina Carvalho & Carpintero, 1986: 611 (gen. nov.; type species: Cafayatina altigena Carvalho & Carpintero, 1986 by original designation); Schuh, 1995: 46 (cat.). syn. nov.
Diagnosis: Microtechnites is recognized by the following combination of characters: macropterous and brachypterous morphs; dark ground colour; moderate distribution of white scale-like setae; eyes sessile; parameres subequal in size; left paramere L-shaped; male endosoma with numerous elongate spicules, often serrate; and, female sclerotized rings widely separated, diagonally orientated. Microtechnites is similar to Halticus in salient features, but is separated from the latter by the elaborate endosomal spicules and the widely separated sclerotized rings.
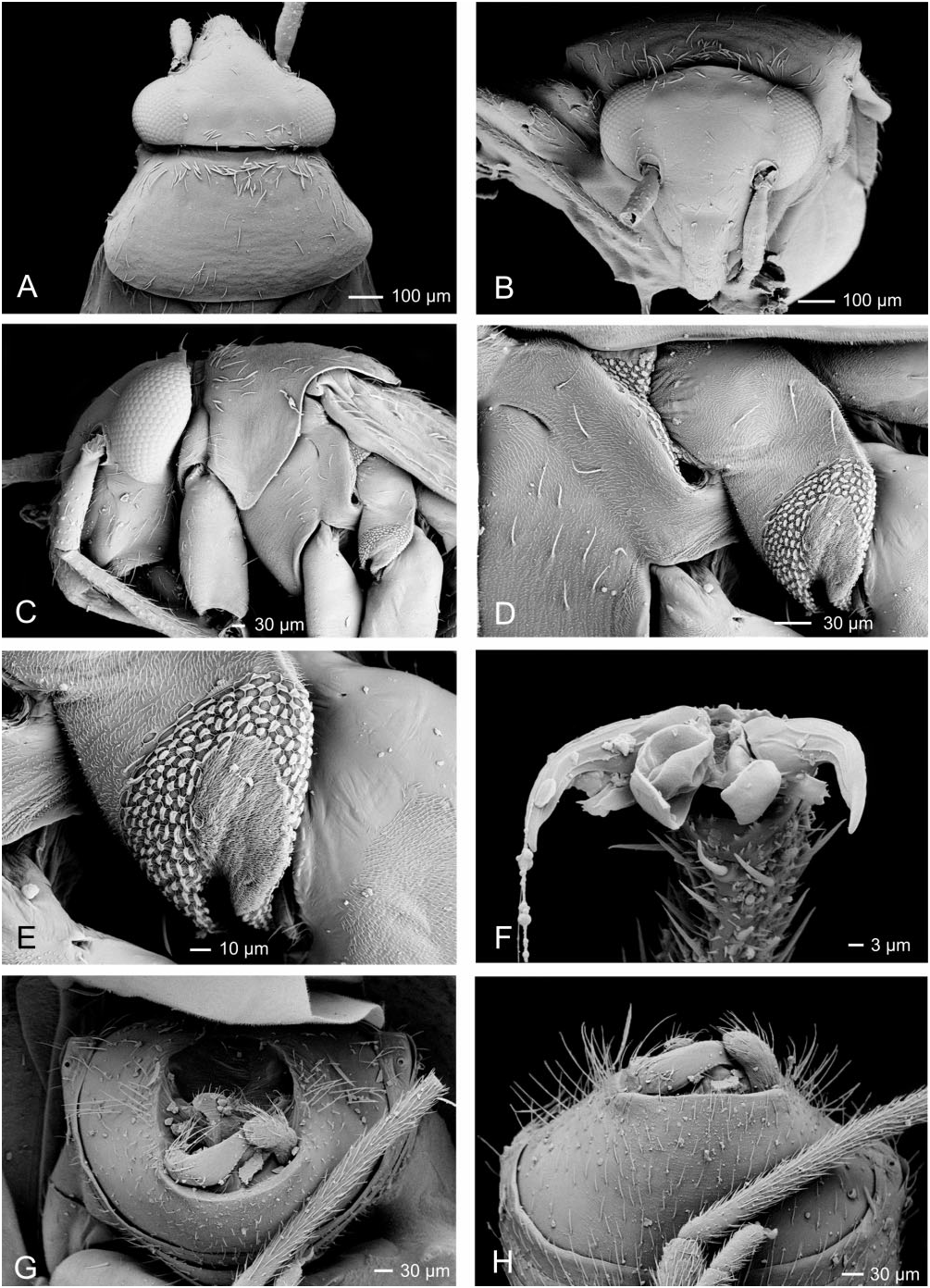
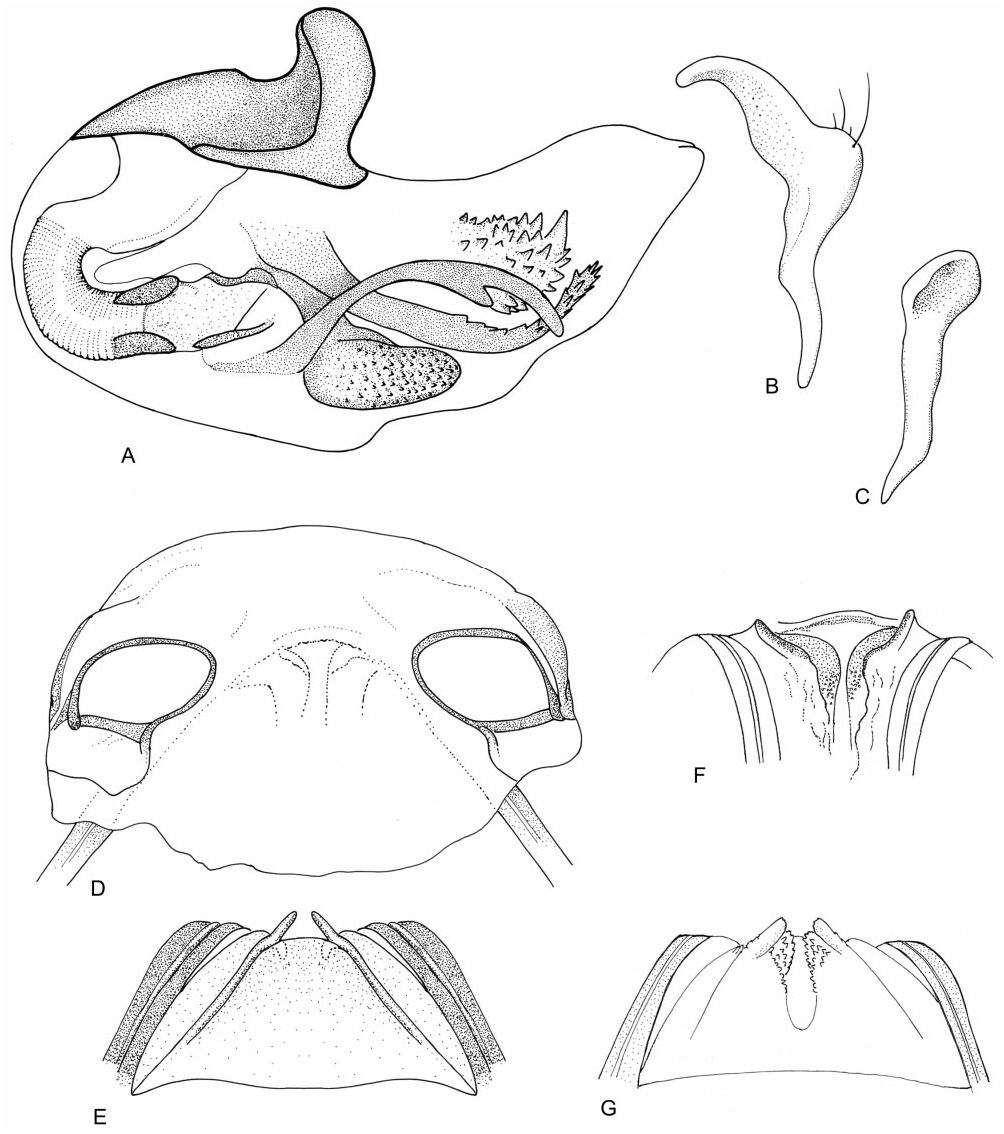
Redescription: Male and female macropterous and brachypterous morphs, sometimes males macropterous only (e.g. M. bractatus ), very small to small, 1.5– 3 mm. Coloration ( Fig. 4 View Figure 4 ): body mostly dark brown with yellow-brown and yellow markings. Surface and vestiture ( Figs 4 View Figure 4 , 37A–H View Figure 37 ): body sparsely clothed in semi-erect simple setae intermixed with decumbent, scale-like setae; legs with semi-erect spines. Head mostly smooth, posterior of pronotum weakly rugulose. Structure: head ( Figs 4 View Figure 4 , 37A–C View Figure 37 ): transverse, elongate in lateral view; posterior margin of vertex sublinear, carinate; head slightly broader than anterior margin of pronotum; frons convex; height of genae approximately equal to height of eye; eyes sessile; maxillary plates sometimes carinate; bucculae narrow; antennae ( Figs 4 View Figure 4 , 37A–C View Figure 37 ): antennal insertion in front of eye; cylindrical, variable length; AI subequal to eye height, weakly swollen; AII- AIV thinner. Labium ( Fig. 37C View Figure 37 ): reaching metacoxae; LI slightly enlarged; thorax ( Figs 4 View Figure 4 , 37A, C–E View Figure 37 ): pronotum trapezoidal, collar present and thin, callosite region weakly delineated, posterior margin convex, humeral angles rounded, in lateral view pronotum of macropterous individuals gently sloping, nearly vertical in brachypterous individuals; mesoscutum not visible in both macropterous and brachypterous individuals; scutellum flat, apically rounded; metathoracic spiracle small uniformly bounded by evaporative bodies, sometimes extending to mesepimeral knob; MTG external efferent system broad, pear-shaped, small, confined to medial third of metespisternum, peritreme oval, medially positioned. Hemelytra ( Fig. 4 View Figure 4 ): macropterous: lateral margins weakly arcuate; embolium widening towards cuneal fracture; clavus broad, claval commissure elongate; median flexion line half length of claval suture; cuneus transverse; membrane with one cell, minor cell obsolete, membrane extending beyond apex of abdomen. Brachypterous: rounded and shell-like, undivided, posterior margins weakly rounded, wholly or partly covering abdominal tergite VI. Legs ( Figs 4 View Figure 4 , 37F View Figure 37 ): metafemora elongate, incrassate; metatibiae elongate; pretarsi with fleshy pulvilli. Abdomen: short, lateral margins parallel, slightly broader in female. Male genitalia ( Figs 37G, H View Figure 37 , 38A–C View Figure 38 ): pygophore short, conical, genital aperture large, posterior margin symmetrical, concave; left paramere L-shaped, sensory lobe small, apophysis broad, apically arcuate; right paramere subequal to left paramere length, spoon shaped; phallotheca simple; ductus seminis short; secondary gonopore with irregular sclerotization; endosoma with several spicules, most spicules elongate, sometimes serrate. Female genitalia ( Fig. 38D–F View Figure 38 ): sclerotized rings moderately sclerotized, widely separated, lateral margins with adjacent portion of DLP weakly upturned; margin of VLP adjacent to rami narrowly sclerotized, entire, without medial flange; posterior wall of bursa copulatrix subtriangular, weakly sclerotized, anteriorly with paired short digitiform processes; vestibulum symmetrical and weakly sclerotized, anteriorly with paired sclerites converging posteriorly, forming a Y-shape.
Diversity and distribution: Microtechnites is restricted to the Western Hemisphere, with most species found in Central and South America. It is the only halticine genus found in the Neotropical region, including its distribution in the Caribbean and the Galapagos Islands ( Henry & Wilson, 2004). Only Mi. bractatus is known in the Nearctic region, being widespread in eastern Canada and the USA and extending as far south as Argentina . Microtechnites chrysolepis is endemic to Hawaii and its distributional isolation is significant. We do not regard this disjunction as necessarily anomalous, as the derivation of Hawaii’s fauna from the Western Hemisphere has been postulated for the orthotyline genus Sarona ( Asquith, 1995) . Included species: Microtechnites altigena ( Carvalho & Carpintero, 1986) * comb. nov. Argentina
Microtechnites bractatus ( Say, 1832) View in CoL * Neotropical and Nearctic regions
= Rhinacloa citri Ashmead, 1887: 155 View in CoL (syn. Reuter, 1909)
= Halticus minutus Uhler, 1890 View in CoL nec Reuter, 1884 (junior primary homonym of Halticus minutus Reuter, 1884 View in CoL )
= Halticus uhleri Giard, 1892:81 (new name for Halticus minutus Uhler – syn. Parshley, 1915)
Microtechnites canus Distant, 1893 View in CoL * (restored status, Henry, 1983: 607) Guatemala; Mexico; Nicaragua
Microtechnites chrysolepis ( Kirkaldy, 1904) View in CoL * Hawaii
Microtechnites inesalti ( Carvalho & Carpintero, 1992) View in CoL * comb. nov. Argentina View in CoL
Microtechnites spegazzinii Berg, 1883 View in CoL * Neotropical region
Biology and host plant associations: Host plants have been recorded only for the ubiquitous Mi. bractatus ; Maldonado (1969) recorded two cucurbit species as host plants, whereas Linnavuori (1984) recorded Mi. bractatus on sweet potato ( Ipomoea batatas ) ( Table 1). This species, which is commonly known as the garden fleahopper, has been recorded as a pest from a broad range of crops ( Henry, 1983; Carpinera, 2001), including solanaceous crops (aubergine, tomato, potato, pepper, tobacco), commercial cucurbits (cucumber, pumpkin, squash) and legumes (alfalfa, beans, clover, cowpeas, peas), as well as beets, cabbage, celery, lettuce, barley, corn, oats, wheat, and cotton. Carpinera (2001) also recorded this species from a great number of weed species, regarded as secondary hosts associated with garden vegetables ( Table 1). Microtechnites bractatus is also thought to be zoophytophagous ( Carpinera, 2001). It has five instars and overwinters as eggs. Eggs are laid in stems and leaf petioles.
Remarks: We have removed Microtechnites from synonymy with Halticus and reinstated it to generic status. In addition, Cafayatina is established as a junior synonym of Microtechnites . Microtechnites is distinguished by its unique male genitalia. In Halticus the aedeagus ends with an elongate scoop-shaped secondary gonopore and the endosoma is vestigial without spicules, whereas in Microtechnites the secondary gonopore opens into a membranous endosoma, which contains several complex spicules and sclerotized serrations. Additionally, the sclerotized rings of Microtechnites are generally smaller and more widely separated, in comparison to Halticus where they are transverse, large, and medially subcontiguous.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Microtechnites
| Tatarnic, Nikolai J. & Cassis, Gerasimos 2012 |
Cafayatina
| Carvalho JCM & Carpintero DL 1986: 611 |
Microtechnites canus
| Henry T 1983: 607 |
Microtechnites
| Cassis G & Gross GF 1995: 186 |
| Carvalho JCM 1958: 13 |
| Reuter OM 1910: 149 |
| Berg C 1883: 73 |