
Chorosomella, HORVATH, 1906
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00770.x |
|
persistent identifier |
https://treatment.plazi.org/id/03E8878D-FFF5-FFF9-5EEC-F95AB058FF79 |
|
treatment provided by |
Marcus |
|
scientific name |
Chorosomella |
| status |
|
CHOROSOMELLA HORVÁTH View in CoL ( FIGS 3 View Figure 3 , 13–14 View Figure 13 View Figure 14 )
Chorosomella Horváth, 1906: 545 View in CoL (gen. nov.; type species: Chorosomella jakowleffi Horváth, 1906 View in CoL by monotypy); Carvalho, 1959: 283 (cat.); Kerzhner,
1962: 385 (tribal placement); Wagner, 1973: 107 (description); Schuh, 1995: 46 (cat.)
Diagnosis: Chorosomella is distinguished from all other halticines by the following combination of characters: gracile body; elongate appendages; rectangulate head; frons with conical projection; vertex with midline sulcus; bucculae narrow; eyes distant from anterior margin of pronotum; AI greatly elongate and thick; labium short, reaching mesocoxae; MTG external efferent system obsolete; pretarsi with pulvilli; single elongate endosomal spicule.
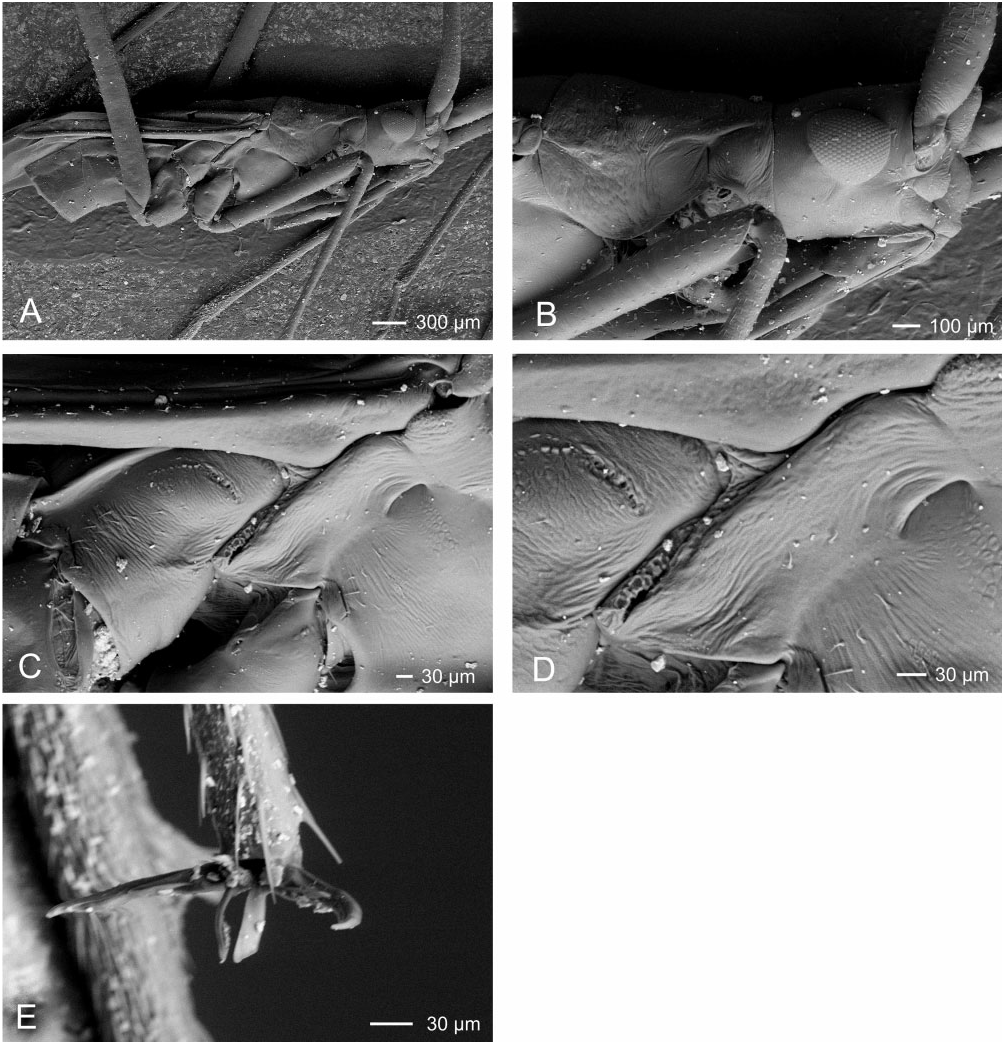
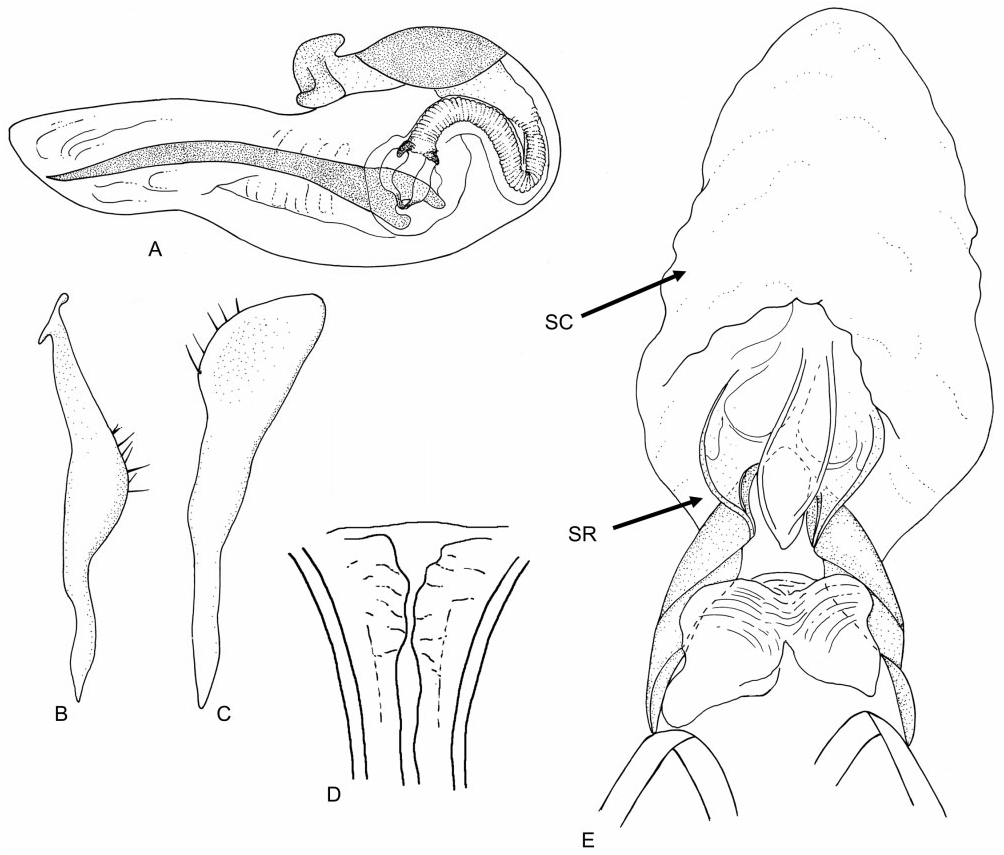
Redescription: Male macropterous, female micropterous. Coloration ( Fig. 3 View Figure 3 ): pale yellow-brown. Surface and vestiture ( Figs 3 View Figure 3 , 13A–D View Figure 13 ): smooth, almost glabrous. Antennae clothed in very short adpressed spinose setae, becoming most dense on AII through AIV; femora with very short, adpressed spinose setae primarily on dorsal surface; tibiae with slightly longer, semi-erect spine-like setae, shorter than leg width. Structure: head ( Figs 3 View Figure 3 , 13A, B View Figure 13 ): rectangulate, elongate, length greater than head height; vertex flat with thin medial longitudinal sulcus, posterior margin not carinate; frons with conical projection, extending beyond clypeus; posterior margin of eyes removed from anterior margin of pronotum; genae height subequal to eye height; mandibular and maxillary plates small; bucculae extremely narrow; antennae ( Figs 3 View Figure 3 , 13A, B View Figure 13 ): inserted at eye midheight; elongate, slender, longer than body; AI thickest, longer than head and pronotum combined; AII approximately twice as long as AI; AIII slightly longer than first; AIV short. Labium ( Fig. 13A, B View Figure 13 ): short, nearly reaching mesocoxae. Thorax ( Figs 3 View Figure 3 , 13A–D View Figure 13 ): pronotum narrow, trapezoidal, posteriorly only slightly wider than anterior margin, nearly flat, collar absent, callosite region distinct, lateral margins rounded, posterior margin medially concave; mesoscutum elongate; scutellum short, apex blunt, rounded; metathoracic spiracle a thin slit, with evaporative bodies along posterior margin, MTG external efferent system obsolete. Hemelytra ( Fig. 3 View Figure 3 ): males macropterous, costal margin concave, becoming very narrow towards apex; membrane with single cell, apex reaches abdominal tergite VII, minor cell obsolete; females micropterous, hemelytra translucent; without cuneal fracture or membrane; only reaching abdominal tergite III. Legs ( Figs 3 View Figure 3 , 13A, B, E View Figure 13 ): very long, cylindrical, thin; pretarsi with fleshy pulvilli. Abdomen ( Fig. 1 View Figure 1 ): elongate in both sexes. Male genitalia ( Fig. 14A–C View Figure 14 ): pygophore elongate, posterior margin laterally with pair of caudally projecting processes; parameres roughly equal in size; left paramere narrow, weakly bent, weakly concave, apex of apophysis bifid; right paramere weakly concave, without apical apophysis; phallotheca elongate, narrow and simple, apically slender; ductus seminis elongate, very thin; secondary gonopore weakly sclerotized, opening into elongate tubular spine-like spicule, reaching apex of phallotheca. Female genitalia ( Fig. 14D View Figure 14 ): sclerotized rings thin, elongate, ribbon-like, very weakly sclerotized. DVP and VLP membranous, without sclerotization; posterior wall of bursa copulatrix entirely membranous, without processes or modifications vestibular region symmetrical. Vestibular region symmetrical, very weakly swollen, with margins wrinkled and membranous, converging anteriorly.
Diversity and distribution: Chorosomella is comprised of two species from central districts of the Palaearctic region, including the Russian Crimea and Kazakhstan.
Included species: Chorosomella horvathi Kiritshenko, 1911 Kazakhstan
Chorosomella jakowleffi Horváth, 1906 View in CoL * Russia
Biology and host plant associations: Chorosomella jakowleffi is known to breed and feed on grasses ( Kerzhner & Yachevskii, 1967; Wagner, 1973; Schuh & Lattin, 1980) ( Table 1).
Remarks: Chorosomella was previously placed in the Mirinae : Stenodemini ( Horváth, 1906) based on superficial similarities, such as the elongate body and appendages, synonymous with many grass-feeding true bugs. Reuter (1910) transferred the genus to the Orthotylini , but Carvalho (1952) subsequently restored it to the Stenodemini . Kerzhner (1962) first moved Chorosomella to the Halticini , whereas Wagner (1973) regarded the halticines as a subfamily, and erected a new tribe for Chorosomella : the Chorosomellini. Most authors follow Kerzhner (1962), who based his decision on various characters he considered typical of the Halticini : L-shaped left paramere, spoon-shaped right paramere, membranous aedeagus with spicula, and tall genae, and supported by their close relationship with graminoids. In examining Chorosomella we find that some of these character evaluations are disputable; compared to other Halticini the left paramere is much less angular; the secondary gonopore is unlike that of any other Halticini , and the genae are considerably shorter. Additionally, the long, gracile body shape, long thin legs, pale coloration, the unique shape of the head, and the structure of the female genitalia, particularly the sclerotized rings, are also unlike all other Halticini .
In our unweighted phylogenetic analysis Chorosomella groups with Dimorphocoris and Compositocoris ( Fig. 1 View Figure 1 ) by several homoplasies, including an overall nonblack coloration (1-0), a slightly elongate head (10-1), and the extreme brachyptery of females, with the wings reduced to wing pads (37-3). However, under implied weighting this relationship dissolves, with Chorosomella now appearing as sister to Plagiotylus in a clade that also includes Strongylocoris . This clade is supported by the lack of metathoracic evaporative area (24-0) and the extreme reduction of the metathoracic scent gland itself (23-0). Based on the current data it is unclear which of these competing topologies more closely reflects the true placement of Chorosomella .
COMPOSITOCORIS SCHWARTZ, SCHUH & TATARNIC
( FIGS 3 View Figure 3 , 15–16 View Figure 15 View Figure 16 )
Compositocoris Schwartz, Schuh & Tatarnic, 2008: 23 View in CoL (gen. nov.; type species: Compositocoris senecionus Schwartz et al., 2008 View in CoL by original designation)
Diagnosis: Compositocoris is recognized by the following combination of characters: strongly sexually dimorphic, males macropterous and elongate, females ovoid and micropterous; bright green coloration; females with apically stellate bristle-like setae; medial margins of first gonapophyses forming vestibular opening symmetrical and weakly sclerotized. This genus is very similar to Dimorphocoris but is distinguished by the shorter genal height, coloration, and the apically stellate setae of the females.
Diversity and distribution: Compositocoris is a monotypic genus, known only from Namaqualand, South Africa.
Included species: Compositocoris senecionus Schwartz et al., 2008 * South Africa
Biology and host plant associations: This species is known from numerous species of asterids belonging to the genera Senecio, Leyesera , and Ursinia (Asteraceae) ( Table 1).
Remarks: Compositocoris is very similar to Dimorphocoris in many aspects, including overall male and female body form, the presence of paddle-like sclerites apical to the secondary gonopore, and the patches of honeycombed texturing on the head and thorax (2-1: the only unambiguous synapomorphy linking these as sister taxa in our phylogeny). Only the bright coloration, shorter head, and stellate bristle-like setae in the female distinguish Compositocoris . A more thorough examination of Dimorphocoris is necessary to identify the range of morphological variation and to determine if Compositocoris is congeneric; this was beyond the scope of this study, as we only had access to a few species of the speciose Dimorphocoris .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Chorosomella
| Tatarnic, Nikolai J. & Cassis, Gerasimos 2012 |
Compositocoris
| Schwartz MD & Schuh RT & Tatarnic N 2008: 23 |
Chorosomella Horváth, 1906: 545
| Carvalho JCM 1959: 283 |
| Horvath G 1906: 545 |