
Cophixalus petrophilus, Hoskin, 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3722.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:22E57934-07E1-4C11-B502-06C5BCB9C93B |
|
persistent identifier |
https://treatment.plazi.org/id/03E887CA-FF82-FF80-FF52-60DB1A84F92D |
|
treatment provided by |
Felipe |
|
scientific name |
Cophixalus petrophilus |
| status |
sp. nov. |
Cophixalus petrophilus View in CoL sp. nov.
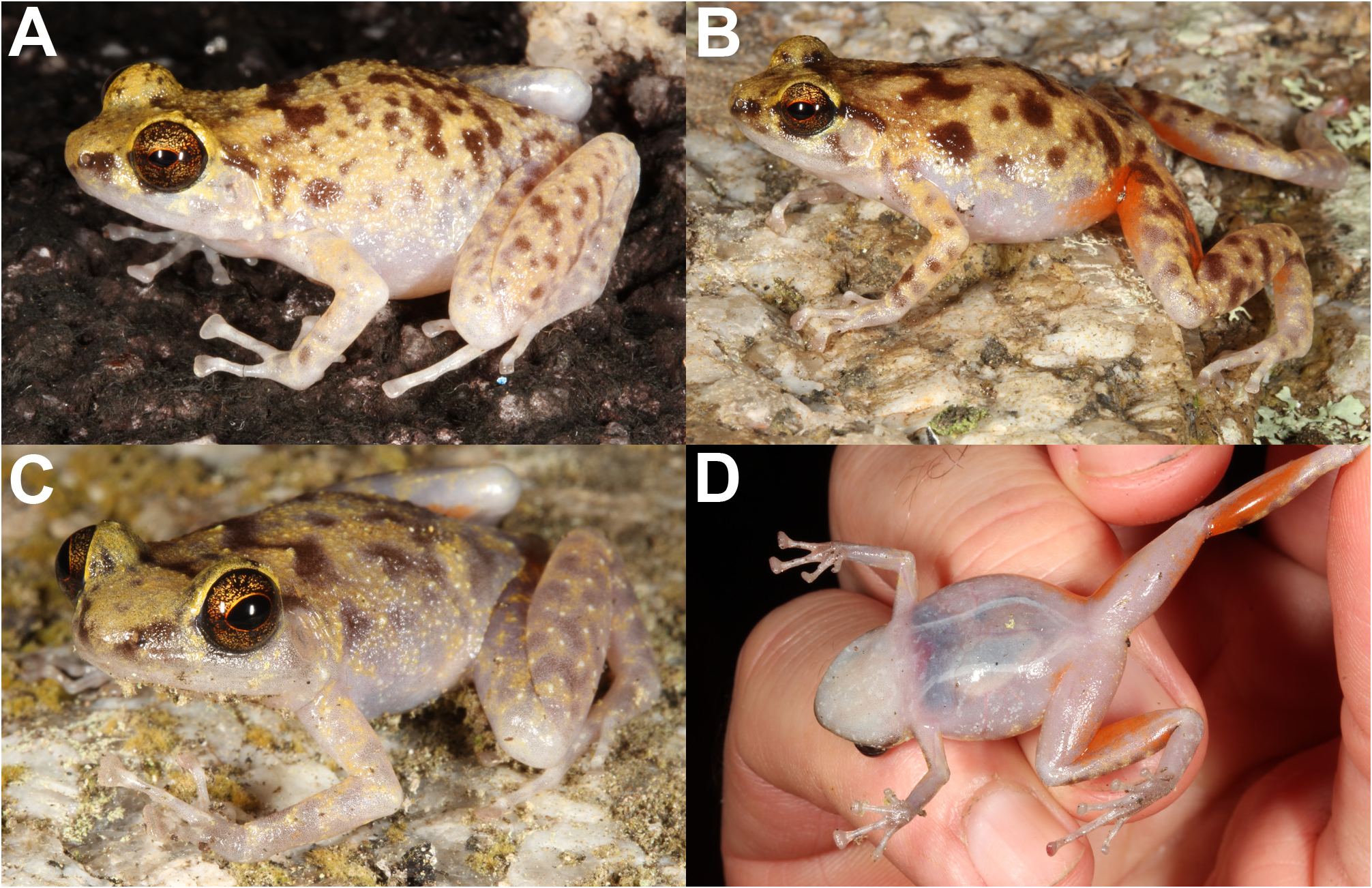
Blotched Boulder-frog
Figs 1 View FIGURE 1 , 2A, 2B View FIGURE 2 , 3 View FIGURE 3
Material examined. Holotype: QMJ92560, male, Melville Range (14°18'54" S, 144°29'54" E; elevation 120 m a.s.l.), Cape Melville, north-east Queensland, C. J. Hoskin, 10 February 2013 GoogleMaps . Paratypes: QMJ92568 (female) , QMJ92569 (male) , QMJ92573 (female) , QMJ92574 (male), Melville Ra. (14°16'44" S, 144°29'22" E; elevation 470 m a.s.l.), north-east Queensland, C. J. Hoskin, 20 March 2013 GoogleMaps .
Diagnosis. Cophixalus petrophilus sp. nov. can be distinguished from all congeners by the following characters: medium body size (SVL 26.4–31.8 mm), blotched dorsal pattern of males and females, and large eyes (ED/SVL = 0.11–0.14). The mating call is also distinct from all Cophixalus , being a medium paced, low-pitched rattle.
Etymology. From the Latin, petrophilus refers to ‘rock-loving’, in recognition of the restriction of this species to boulder-field habitat. The species epithet is treated as a noun in apposition.
Measurements of holotype (mm): SVL 28.3; HLL 14.1; FLL 7.4; HW 10.0; HL 6.9; HD 3.4; ED 3.7; EN 1.9; IN 1.9; TYMP 2.2; 3DW 1.5; 3FL 4.3; 4DW 1.0; 4TL 6.7.
Description of type series. Data presented as range followed by mean in brackets. Adult measurements (mm): SVL 26.4–31.8 (29.2); HLL 13.3–15.4 (14.4); FLL 6.8–8.7 (7.5); HW 9.8–11.8 (10.8); HL 6.9–8.6 (7.7); HD 3.4–4.8 (4.1); ED 3.2–4.2 (3.7); EN 1.9–2.7 (2.3); IN 1.7–2.3 (2.0); TYMP 1.7–2.2 (1.9); 3DW 1.4–1.9 (1.6); 3FL 4.1–5.1 (4.6); 4DW 0.8–1.2 (1.0); 4TL 6.3–7.7 (6.9). Adult proportions: Table 1. Comparison of sexes: Females are larger than males (female SVL 31.2–31.8 mm, male SVL 26.4–28.3 mm) but no major differences in proportions were detected between the sexes ( Table 1). Head: Narrower than body, triangular in dorsal view; snout truncated at the nares, blunt to projecting in profile; canthus rostralis rounded to angular, loreal region steep; nares much closer to tip of snout than to eye, nares anterolateral on tip of snout; eyes large; eye diameter greater than eye to naris distance; internarial distance about equal to distance from eye to naris; tympanum large (approximately half diameter of eye), indistinct to moderately distinct beneath overlying skin, bordered dorsally by supra-tympanic fold. Body: Oval. Limbs: Hindlimbs and forearms relatively long; fingers and toes unwebbed; relative finger length 3>4>2>1; fingers 2, 3 and 4 long and slender with large, truncated discs; first finger short (about one-third to half length of second) with small and rounded disc; broad, low, rounded outer palmar tubercles; subarticular tubercles low, moderately prominent; relative length of toes 4>3>2>5>1 (2 and 5 about equal length), toes long and slender, toe 4 very long; toe discs moderately truncated, discs smaller on toes 1 and 5; low, rounded inner metatarsal tubercle, no outer metatarsal tubercle; subarticular tubercles low and rounded, moderately prominent; discs on fingers larger than discs on toes. Skin: Dorsal surfaces smooth, with scattered fine tubercles; ventral surface smooth; indistinct supra-tympanic fold. Colour pattern in preservative ( Fig. 3 View FIGURE 3 ): All dorsal surfaces cream or creamy brown with darker brown blotches. Blotching includes a prominent W-shaped mark on the shoulders, a dark bar across the top of the head between the eyes, and irregular blotching over the lower back and limbs. Dark band around wrists; tops of hands white; top of finger pads grey. Pale lumbar ocelli visible; particularly prominent on the darker males. Greyish triangle on top of snout; eyelids light to dark grey. Dorsal blotching fades out on flanks. Ventral surfaces uniformly pale (white or cream) except for grey shading on undersides of hands and feet. Dark band from nare to above tympanum, broken by eye; in some specimens, rather than a dark band, much of the loreal region is dark. Tympanum pale with light brown center. Iris dark but heavily flecked with silver. Groin and posterior thigh finely mottled and smudged brown and grey; the orange seen in life in the groin, posterior thigh and on the ‘calf’ appears as white or soft orange areas in preserved specimens.
Measurements of live individuals. The holotype was measured in the field: SVL 27.1 mm, WT 1.85 g.
Colour pattern in life ( Figs 1 View FIGURE 1 , 2A, 2B View FIGURE 2 ). Dorsal surfaces of males creamy yellow with dark brown blotching; colours and markings paler in females. Dorsal markings include a prominent dark W-shaped mark on the shoulders, a dark bar across the top of the head between the eyes, irregular dark blotching over the back, dark or grey blotching or mottling on the upper flanks and limbs, and indistinct yellow lumbar ocelli. Top of hands, feet and digits white or pale grey; top of pads grey. Yellow triangle on top of snout and eyelids. Dark band from nare, through eye, and over tympanum. Tympanum pale. Dark horizontal line through iris, similar depth and colour to pupil, bordered above and below by red; upper and lower iris black but heavily flecked with gold. Dorsal blotching and yellow background colour fades out mid-flank to unmarked grey or white lower flank. Ventral surfaces uniformly white or pinkish grey; throat white; internal organs (heart, liver, gut) opaquely visible; two longitudinal white bands visible in males, from near forearm insertion to near hindlimb insertion; undersides of hands and feet grey ( Fig. 1D View FIGURE 1 ). Groin and hidden parts of hindlimbs (posterior thigh, ‘calf’, top of feet) orange; slightly paler in females.
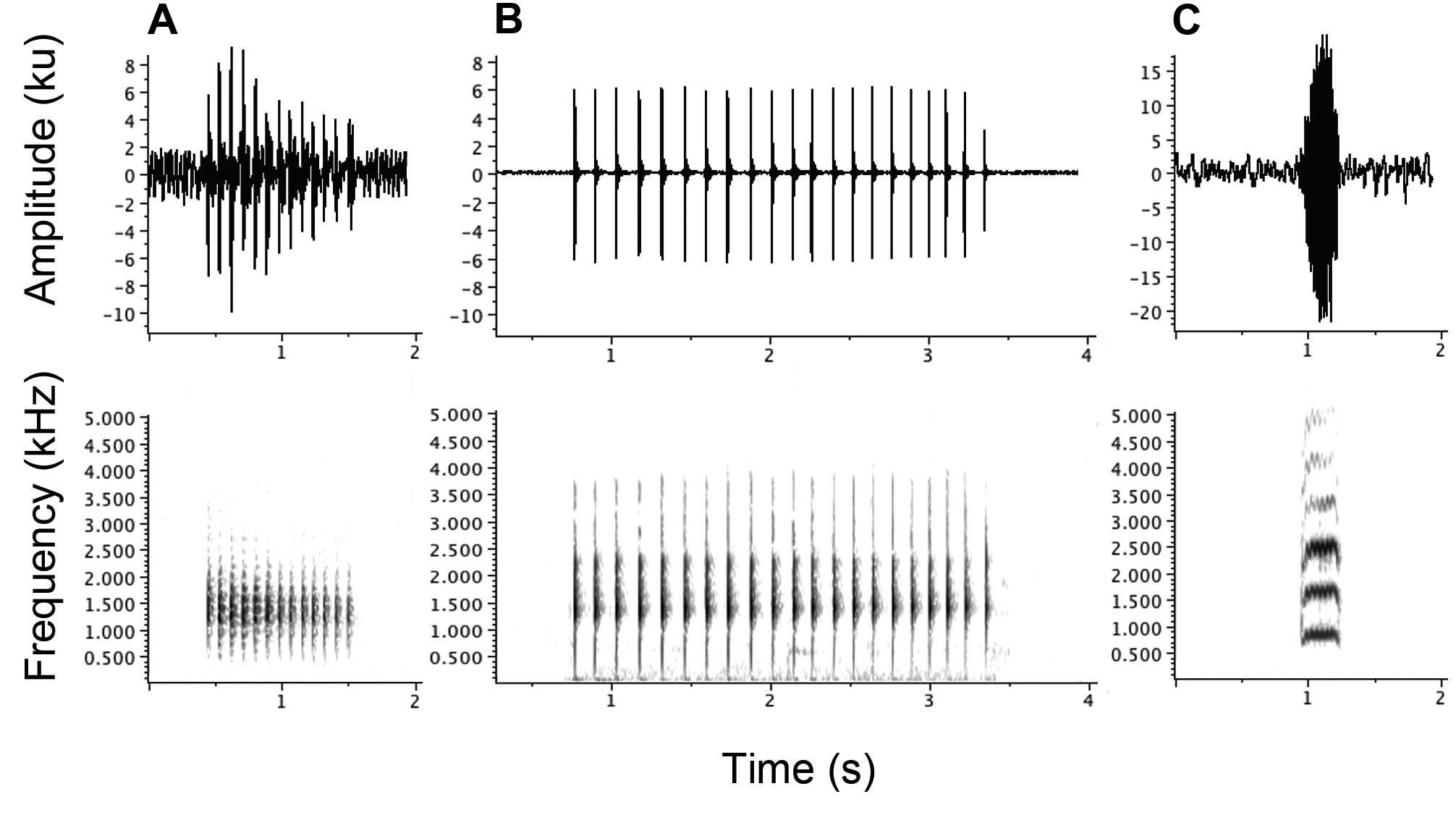
Call. Calls were only obtained for a single individual (holotype, QMJ92560). The mating call of C. petrophilus sp. nov. is a fairly short, medium-paced, low-pitched rattle of the following characteristics (mean followed by range in brackets): duration 1.00 s (0.92–1.07), notes per call 12 (11–13), note rate 12.0 notes/s (11.7–12.2), dominant frequency 1.30 kHz (1.27–1.32) ( Table 1). Calls are uttered at an interval of approximately 3.5 seconds (3.3–3.9). Air temperature at the time of recording was approximately 25ºC. Figure 4A View FIGURE 4 displays a single representative call .
Comparison. Assigned to Cophixalus based on external morphology and genetics (see first paragraph of ‘Systematics’). Distinguished from all congeners by medium body size (male SVL: 26.4–28.3 mm, female SVL 31.2–31.8 mm), large and truncated finger pads, blotched colour pattern of males and females, large eyes (ED/SVL = 0.11–0.14), and low-pitched rattling call. Most closely resembles the other boulder species due to larger size, long fingers, and large, obviously truncated finger pads. The new species differs from these four species as follows. Cophixalus petrophilus sp. nov. is distinguished from C. kulakula by its smaller size (SVL 26.4–31.8 mm vs 39.8– 48.0 mm) and blotched dorsal pattern. Distinguished from C. pakayakulangun by smaller size (SVL 26.4–31.8 mm vs 42.2–52.9 mm), orange groins, and blotched dorsal pattern of males and females. Cophixalus petrophilus sp. nov. is distinguished from C. saxatilis by its smaller size (male SVL 26.4–28.3 mm vs 30.0–35.0 mm; female SVL 31.2–31.8 mm vs 39.9–45.1 mm), larger eyes (ED/SVL 0.11–0.14 vs 0.08–0.11), and larger tympanum, particularly in males (TYMP/SVL 0.063 –0.076 vs 0.049 –0.059) ( Table 1; Fig. 2 View FIGURE 2 ). Additionally, C. petrophilus sp. nov. males are more heavily blotched than male C. saxatilis , and C. petrophilus sp. nov. females are blotched dark on creamy yellow whereas C. saxatilis females are fairly immaculate and bright yellow ( Fig. 2 View FIGURE 2 ). Further, the call of C. petrophilus sp. nov. is a fairly short, medium-paced rattle, whereas that of C. saxatilis is a slow tap ( Table 1; Fig. 4 View FIGURE 4 ). Cophixalus petrophilus sp. nov. co-occurs with C. zweifeli but is readily distinguished from that species by its smaller size (SVL 26.4–31.8 mm vs 36.3–48.9 mm), longer hindlimbs (HLL/SVL 0.49–0.50 vs 0.45–0.49), wider head (HW/SVL 0.35–0.38 vs 0.31–0.37), longer head (HL/SVL 0.24–0.28 vs 0.22–0.23), and larger eyes (ED/SVL 0.11–0.14 vs 0.09–0.10) ( Table 1; Fig. 2 View FIGURE 2 ). Additionally, C. petrophilus sp. nov. has a blotched dorsal surface whereas the dorsal surface of C. zweifeli is immaculate ( Fig. 2 View FIGURE 2 ). The calls of C. petrophilus sp. nov. and C. zweifeli are very different, with that of the former being a low-pitched rattle and that of the latter being a high-pitched ‘yelp’ ( Table 1; Fig. 4 View FIGURE 4 ). Cophixalus petrophilus sp. nov. is readily distinguished from all Australian frogs (except the larger Cophixalus species ) by having long slender fingers with large, truncated finger pads.
Genetics. There is substantial sequence divergence (for 963 bp 12S and 16S rRNA) between C. petrophilus sp. nov. and all other Australian Cophixalus (8.1–13.6%) (Hoskin, unpub. data). Phylogenetic analyses suggest that C. petrophilus sp. nov. is most closely related to C. zweifeli and C. infacetus , with sequence divergence between C. petrophilus sp. nov. and these two species being 8.1% and 8.2%, respectively (Hoskin, unpub. data).
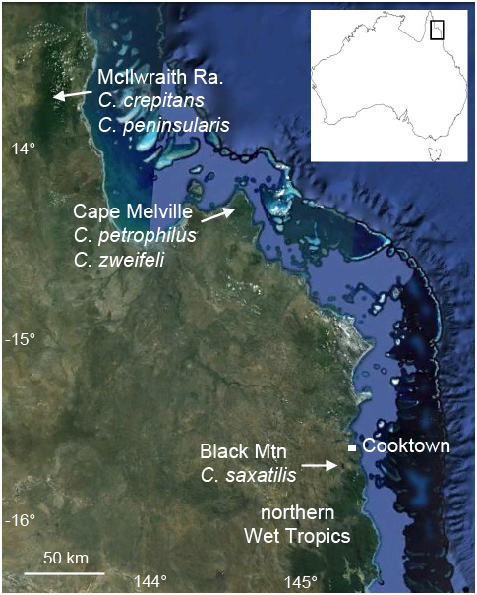
Distribution. Known only from granite boulder-fields of the Melville Range, Cape Melville, north-east Australia ( Fig. 5 View FIGURE 5 ). Cophixalus petrophilus sp. nov. has been found at two sites, one at 120 m elevation at the southern end of Melville Range (14°18'54" S, 144°29'54" E) and the other at 470 m elevation in the central uplands of the range (14°16'44" S, 144°29'22" E). These sites are approximately 4 km straight-line distance apart. Cape Melville is dominated by extensive boulder-fields ( Fig. 6A View FIGURE 6 ), and the species is probably widely distributed through these.
Habitat and habits. Cophixalus petrophilus sp. nov. inhabits deeply piled granite boulder habitat that is largely devoid of vegetation ( Fig. 6A View FIGURE 6 ). At both sites the species was found at night sitting on surface boulders of very deeply piled boulder-field. Exploration below the surface at these sites revealed a deep labyrinth of boulders with no sign of soil or surface water to the depths that could be safely explored. Humidity was very high within the boulder-fields, the rock crevices were moist in places, and water flowed over the rock surfaces during rain and deep into the boulder-field. Indeed during heavy rain, and for a short period after, a loud sound of flowing water could be heard coming from the boulder-fields. Conditions were fairly dry during the survey of the lowland site in February 2013 and only a single individual (the holotype) was found when he called following a brief shower at 5 am. No individuals were found during the preceding night despite extensive searches of the boulder-field and surrounding rainforest. In contrast, conditions were moister during the survey of the upland site in March 2013 and four individuals (two males and two females) were found in a relatively small area of boulder-field (the rock patch in the top left of Figure 6B View FIGURE 6 ). No males were heard calling at this site and the frogs were found at night sitting on surface rocks or moving towards the surface from the depths of the boulder-field. The lack of calling activity at this time of the year conforms to the general pattern in Australian Cophixalus frogs of calling through the first twothirds of the wet season (approximately Sept.–Feb.) but not in the latter part of the wet season ( Hoskin & Aland 2011).
The holotype was calling from a rock crevice about two boulder ‘layers’ below the surface. The crevice was moist and mossy compared to the surrounding rock surfaces. The male was wedged in the crevice, facing out, calling at regular intervals. Cophixalus petrophilus sp. nov. is likely a terrestrial breeder like all other Australian microhylid frogs ( Hoskin 2004; Hoskin & Aland 2011), and it is likely that the male was calling from the nest site. In a similar species, C. saxatilis , in which males also call from exposed boulder-fields, an egg clutch has been found in a rock crevice attended by a male ( Hoskin 2004). No other breeding information is known for C. petrophilus sp. nov.. Other frogs found in the vicinity of both sites were C. zweifeli and Litoria andiirrmalin McDonald, 1997 , although neither of these species was found in micro-sympatry with C. petrophilus sp. nov.. While all C. petrophilus sp. nov. were found in exposed boulder-field largely devoid of vegetation, the other two species were found nearby in boulder-field abutting rainforest or boulder habitat under a rainforest canopy. Other vertebrates found at the C. petrophilus sp. nov. sites were the gecko Gehyra dubia ( Macleay 1877) at night at both sites, and, during the day, the skink Cryptoblepharus fuhni Covacevich & Ingram 1978 at the lowland site and an undescribed Saproscincus skink (Hoskin, in press) at the upland site.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |