
Otiorhynchus
|
publication ID |
https://doi.org/10.11646/zootaxa.4108.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B802F2B1-944E-4B84-A856-8091E60D88FC |
|
DOI |
https://doi.org/10.5281/zenodo.6062740 |
|
persistent identifier |
https://treatment.plazi.org/id/03EB857D-FFD9-AD29-0592-FEDB42DB126D |
|
treatment provided by |
Plazi |
|
scientific name |
Otiorhynchus |
| status |
|
Otiorhynchus View in CoL (s. str.) tenebricosus (Herbst, 1784) complex: Otiorhynchus clavipes (Bonsdorff, 1785)
Material examined: 6 larvae ( Fig. 213 View FIGURES 211 – 219 ). JKI, Braunschweig, climate chamber, breeding in flowerpots with Euonymus fortunei , 19.12.2013, 2 ex.; 24.01.2014, 1 ex.; dto., breeding with Syringa vulgaris , 13.11.2013, 2 ex.; 24.01.2014, 1 ex.
Remarks about breeding and development. All larvae were obtained from 1 breeding attempt in 2013. Origin of the adults is the “Ziegenberg” in Paderborn-Wewer (Northrhine-Westphalia, Germany), 24.06.2013. It is a beech forest on limestone with many ash trees and other woody plant species in low altitude ( 120–150 m). In the JKI 4 weevil specimens ( 3 females, 1 male) were released in 4 flowerpots with Euonymus fortunei and 1 with Syringa vulgaris on 27.06.2013. The first larvae were seen on 0 4.10.2014 and mature larvae on 13.11.2013.
The Ziegenberg, located in the hill country, is only 45 km west of the border to Niedersachsen, from where this taxon is absent. The Teutoburg Forest is regarded to be the northeastern distribution limit of Otiorhynchus clavipes , and this taxon is replaced in the entire northern part of Germany, i.e. the plains with predominantly sandy soils, by O. lugdunensis . In the neighbouring eastern hill countries including the whole eastern part of Germany, only O. fuscipes is present ( Dieckmann 1980).
The development of Otiorhynchus clavipes is only reported by Ibbotson & Edwards (1954) from strawberry fields at Cheddar in Southwest England, where this species occurred as pest in a small area. Hoffmann (1958: 1733) regarded these weevils as O. lugdunensis , but this is doubtful, as Cheddar is not situated in the sandy plains, but close to limestone forests of low altitudes, where O. clavipes was also found in Northrhine-Westphalia.
According to Ibbotson & Edwards (1954), the life-cycle of Otiorhynchus clavipes is completed within 12 or 18 months depending on the oviposition period. The adult weevils appear in two waves. One part pupates in autumn, overwinters in the pupal chamber and emerges in large numbers from April to May, the other part pupates from mid-May to July, and adults emerge from mid-June to end of August. For pupation larvae move deeper in the soil and pupate between 15 and 20 cm below soil level.
From the data of Ibbotson & Edwards (1954) and our observations on the development of the larva of the closely related Otiorhynchus lugdunensis the statement that the development time of O. clavipes should last 12 or 18 months is not fully plausible. Hence, the following modification of the life-cycle is proposed here: early emerging weevils lay eggs in spring and early summer, larvae develop during spring and summer, pupate and hatch in autumn, overwinter in the pupal cell and emerge again early in the season. And, as usual in many other Otiorhynchus species, specimens that lay eggs in the second half of the summer cannot pupate before winter comes, overwinter as larva and complete their development during spring and early summer, pupate between mid- May and July and hatch from the soil, when the pupa has developed into the adult stage in summer. Then the newly hatched weevils oviposit, and larvae overwinter again.
In this species there is the same phenomenon as e.g. in Otiorhynchus indefinitus or O. raucus , namely that weevils hatch in late summer or early autumn from the pupa, remain soft-bodied in their pupal chamber during the whole winter, and emerge on the soil surface in spring with their still not sclerotized body. But opposite to all other species, the subpopulation with pupation in late spring to early summer produces adults that hatch short-term after pupation from their pupal chamber and sclerotize immediately: apparently sclerotization is strongly temperaturedependent. The proportion of soft-bodied weevils is 100% early in April and low or 0 in the second half of the early emergence period indicating that emergence has stopped and sclerotization is completed. During the second emergence period in summer the proportion of soft-bodied adults is always low indicating a more continuous hatching and a more rapid sclerotization during summer.
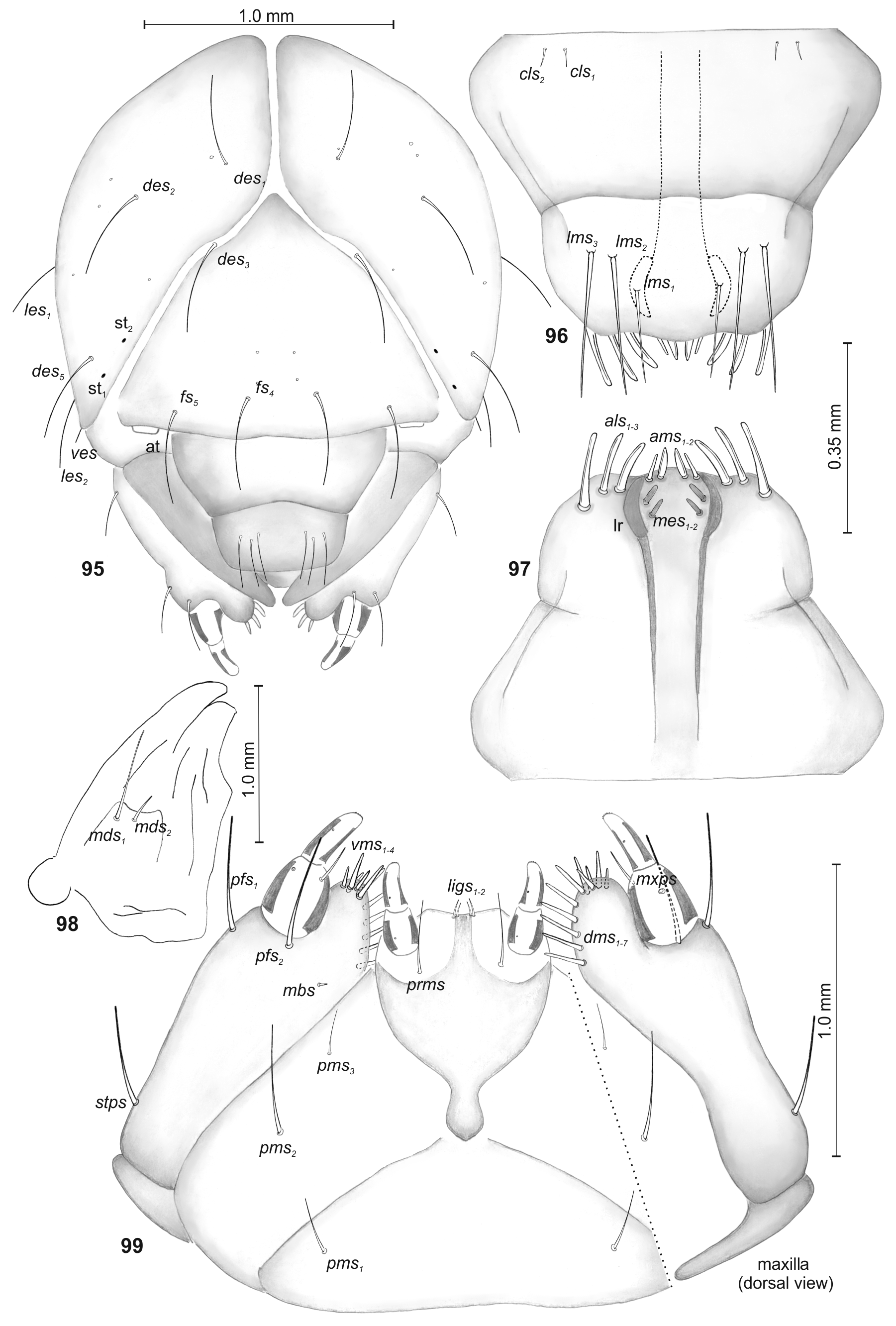
Description ( Figs. 89–99 View FIGURES 89 – 94 View FIGURES 95 – 99 ).
Coloration. Head yellow; all thoracic and abdominal segments yellow; cuticle almost smooth.
Body moderately elongated ( Fig. 89 View FIGURES 89 – 94 ). Chaetotaxy: Setae different in length, filiform, brown. Thorax. Prothorax with 4 long and 4 medium long to short prns; and 2 ps, different in length. Mesothorax with 1 medium long prs; 4 pds (ordered: 2 short and 2 long); 1 long and 1 very short as; 1 long eps; and 1 long ps. Chaetotaxy of meso- and metathorax similar. Each pedal area of thoracic segments well isolated, with 5 pda, different in length. Each thoracic segment with 1 short eus ( Fig. 90 View FIGURES 89 – 94 ). Abdomen. Abdominal segments I–VIII with 1 short prs; 5 pds (ordered: 2 short, 1 long, 1 short and 1 long); 1 medium long and 1 very short sps [abd. seg. VIII only with 1 very short sps]; 2 eps, almost equal in length; 1 long and 1 short ps; 1 long lsts; and 2 eus ( Figs. 91, 93, 94 View FIGURES 89 – 94 ). Abdominal segment IX with 3 ds, different in length; 1 long and 1 very short ps; and 2 sts, equal in length ( Figs. 92–94 View FIGURES 89 – 94 ). Lateral lobes of abdominal segment X with 2–3 minute ts each.
Head subglobose (slightly narrowed) ( Fig. 95 View FIGURES 95 – 99 ). Head capsule with 4 long des, des 1 and des 2 placed on central part of epicranium, des 3 located on frontal suture, des 5 located anterolaterally; 2 long fs, fs 4 placed anteromedially, fs 5 located near to antenna; 2 long les; and 1 very short ves. Postepicranial area with group of 5 pairs of sensilla, frons with 2 pairs of sensilla placed medially. Stemmata (2 pairs) feebly visible. Antennal segment membranous, bearing 1 conical sensorium and 2–3 filiform sensilla. Clypeus 2.4 times as wide as long with 2 cls, equal in length, placed posterolaterally ( Fig. 96 View FIGURES 95 – 99 ). Labrum about 1.8 times as wide as long with 3 straight lms of different length, placed medially or mediolaterally; lms 1 shorter than other setae, all lms exceeding the outline of the labrum; the anterior margin of labrum double sinuate ( Fig. 96 View FIGURES 95 – 99 ). Epipharynx with 3 finger-like als, different in length; 2 ams, different in length; and 2 very short mes, labral rods (lr) short, reniform, strong convergent ( Fig. 97 View FIGURES 95 – 99 ). Mandible ( Fig. 98 View FIGURES 95 – 99 ) bifid, teeth almost of equal height; with 2 mds, different in length; internal edge with a triangular tooth. Maxilla ( Fig. 88 View FIGURES 84 – 88 ) with 1 very long stps and 2 very long pfs, placed ventrolaterally; 1 very short mbs, situated ventrally. Mala with 7 dms, different in length and 4 straight vms, different in length. Maxillary palpi with two palpomeres almost equal in length; basal palpomere with 1 mxps; distal palpomere with a group of 6 conical, cuticular processes apically; each palpomere with a sensillum. Praelabium heart-shaped ( Fig. 99 View FIGURES 95 – 99 ), with 2 very short ligs and 1 long prms. Labial palpi with two palpomeres, relatively elongated; both palpomeres almost equal in length; praemental sclerite wide, well visible. Postlabium with 3 pms, different in length; pms 2 very long, 2 times as long as pms 1 and pms 3 ( Fig. 99 View FIGURES 95 – 99 ).
Differential diagnosis. See “Key to larvae of selected Otiorhynchus species” and Tables 1, 2.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Entiminae |