
Usnea taylorii
|
publication ID |
https://doi.org/10.1016/j.phytochem.2019.04.019 |
|
DOI |
https://doi.org/10.5281/zenodo.10580653 |
|
persistent identifier |
https://treatment.plazi.org/id/03EC9B2A-5101-BB43-FF84-1702FA98FBF7 |
|
treatment provided by |
Felipe |
|
scientific name |
Usnea taylorii |
| status |
|
2.1. Morphological and chemical characterisation of Usnea taylorii View in CoL
2.1.1. Description of Usnea taylorii morphology
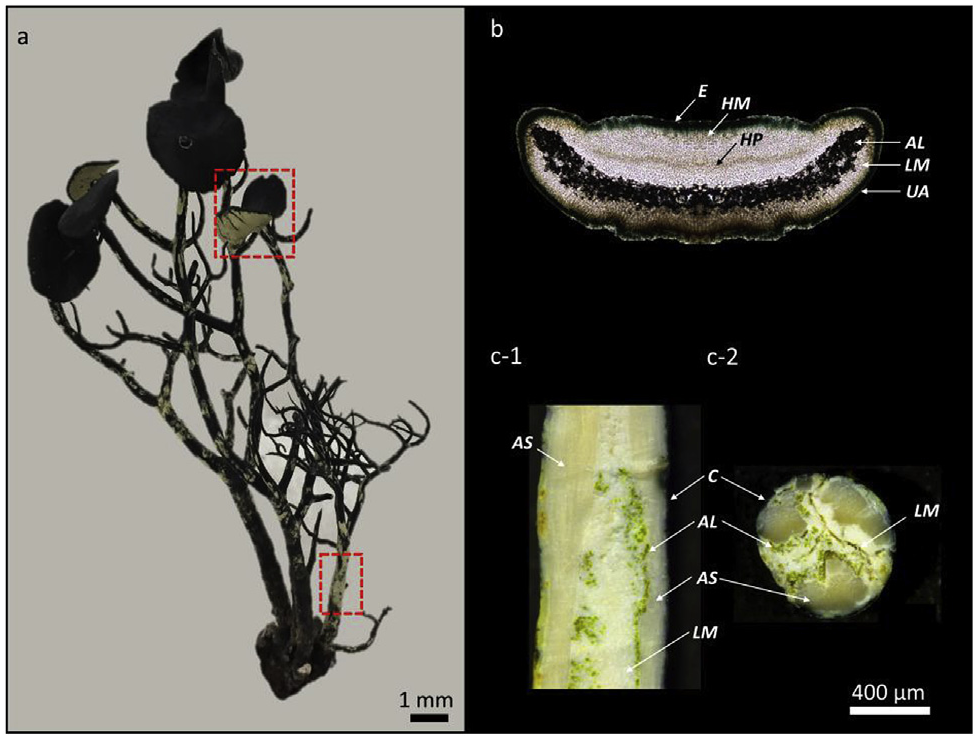
Microscopic analysis of the fruticose lichen Usnea taylorii reveals a specific morphology that deserves a detailed description to better understand the location of metabolites ( Fig. 1a; a View Fig complete morphological description of the lichen is available in Text S1). Apothecia (the reproductive parts of the lichen) are located at the apex of the Usnea branches and are characterised by their large diameter ( 2–17 mm) and their jet-black pigmented disc. In cross-section, underneath the black epithecium, asci containing spores are present in the hymenium, covering the algal layer ( Fig. 1b View Fig ). The thallus in cross-section shows a very thin cortex, protecting a discontinuous algal layer. The central axis of U.
taylorii is divided into several smaller axial strands by the protruding medullae, which are surrounded by some algae. Between these axial strands, lax medulla is observed ( Fig. 1c View Fig ).
2.1.2. Chemical characterisation of Usnea taylorii
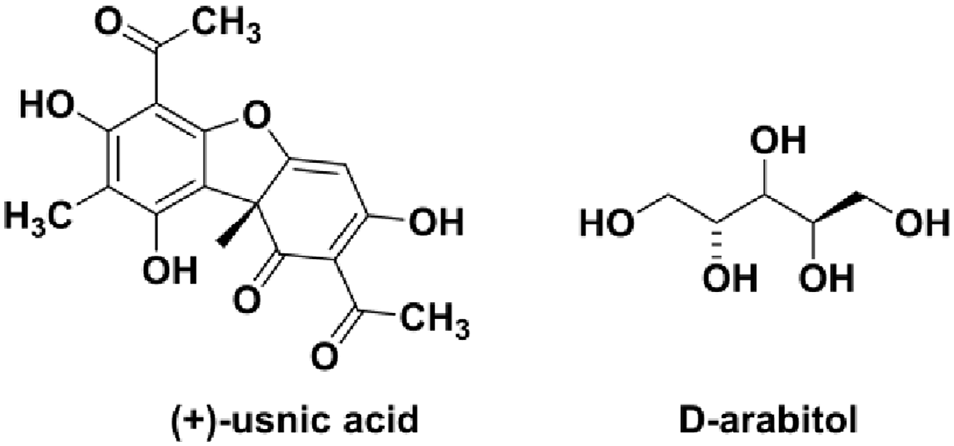
The extraction yields with acetone ranged between 0.5 and 1.0% of specialised metabolites for the six replicates of dried lichen materials with a mean value of 0.8 ± 0.2%. Preliminary experiments showed that more than 95% of the specialised compounds were extracted from the lichen U. taylorii by rinsing intact air-dried thalli in acetone. Thalli had only one or two main specialised metabolites. The concentration of usnic acid ranged from 2.5 to 5.4 mg g −1 dry mass ( DM) of lichen, with a mean value ± s.d. of 4.1 ± 1.1 mg g −1 DM (n = 6). Sugar and polyol profiling was also performed. A low diversity was observed, with only four metabolites quantified. Arabitol was the most important polyol, reaching 138.4 ± 25.8 mg g −1 DM (mean value ± s.d., Table S1 View Table 1 , Fig. 2 View Fig ).
Preliminary LDI-MS analysis of an acetone extract of Usnea taylorii confirmed that the (+)-usnic acid was the only specialised metabolite detected through its deprotonated molecule ( m/z 343) along with a fragment ion at m/z 329 ([M-Me] -) ( Fig. S1 View Fig ). Usnic acid was consequently imaged through the m/z 343 ion. D-arabitol, the major polyol quantified by GC, was also imaged by mass spectrometry. However, because D-arabitol does not absorb at the wavelength of the laser used for LDI-MSI, a MALDI matrix solution was sprayed on U. taylorii slices and images were obtained using MALDI-MSI (through its sodium adduct observed at m/z 175).
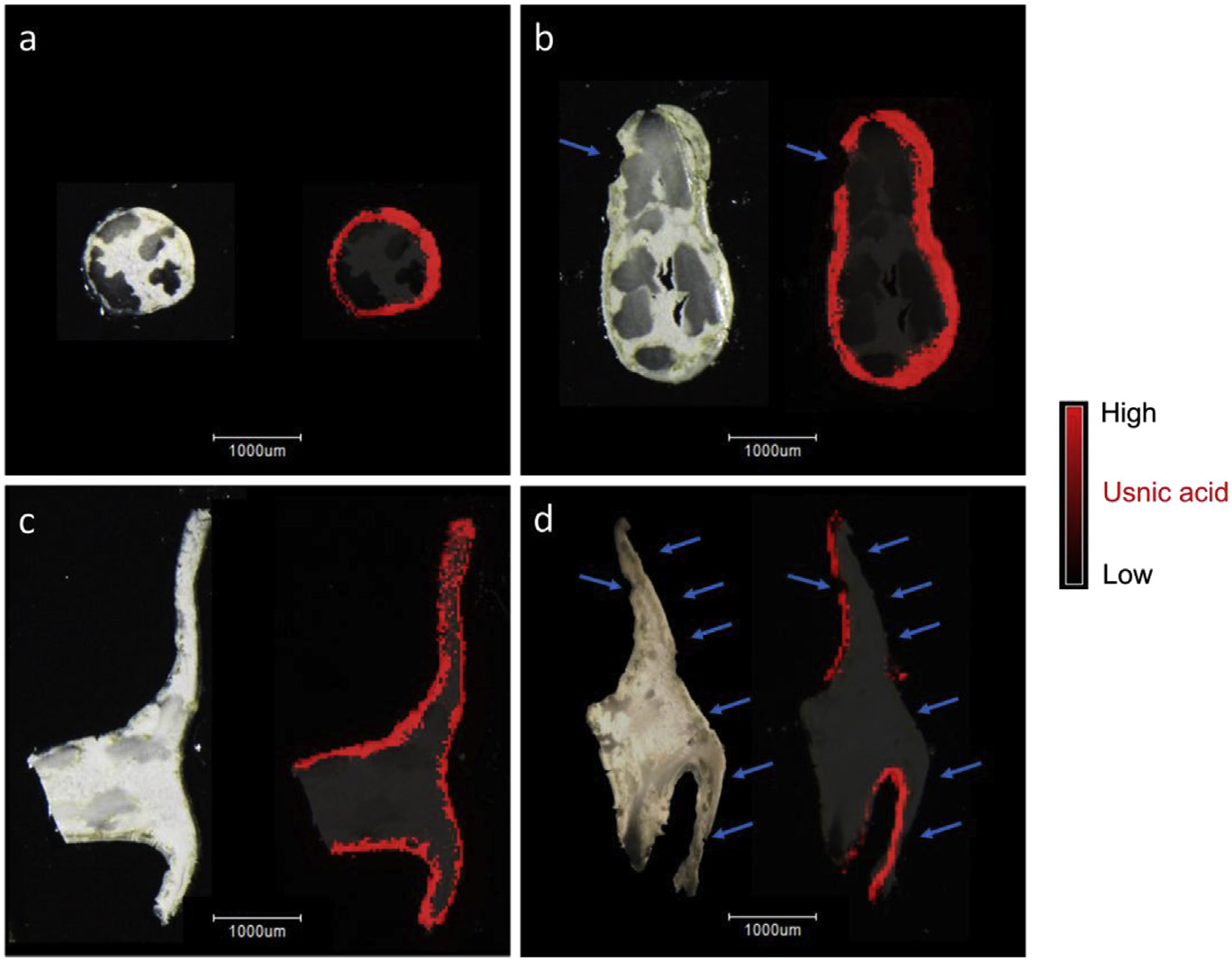
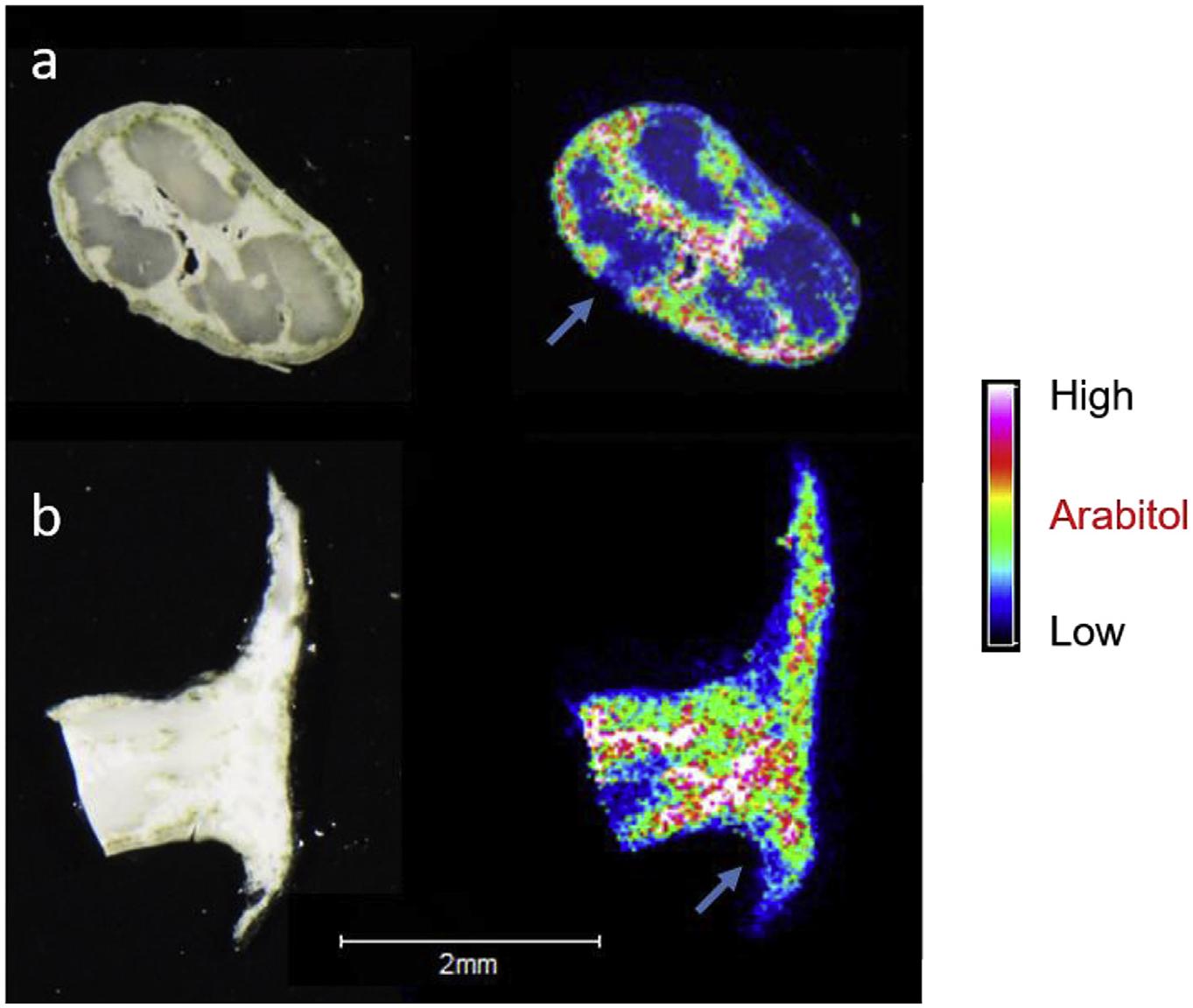
In situ LDI-MSI experiments applied to a slice of Usnea taylorii 's branch revealed that usnic acid was located in the peripheral layer of the thallus, i.e., in the cortex and in the uppermost parts of the medulla including the photobiont layer ( Fig. 3a View Fig ). Likewise, usnic acid was allocated to the external layers of the apothecium (epithecium, hymenium and the underside of the apothecium) ( Fig. 3c View Fig ). D-arabitol was present mainly in the lax medulla, the cortex and the algal layer ( Fig. 4a View Fig ), but occurred in lower intensities in the axial strands of the lichen branches. In apothecia, the signal corresponding to D-arabitol was stronger in the layer containing the lax medulla and algae than in the external layers of apothecium (epithecium and hymenium) ( Fig. 4b View Fig ). To perform a comparison between lichen parts eaten by snails or not, some branches and apothecia were sliced and analysed by LDI-MSI. Snails consumed parts of the cortex, the algal layer and the lax medulla of the branches. In apothecia, epithecium, hymenium as well as the underside of the apothecium were grazed. All these tissues contain usnic acid ( Fig. 3b and d View Fig ) and D-arabitol ( Fig. 4 View Fig ).
| DM |
Dominion Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |