
Chazara briseis
|
publication ID |
https://doi.org/10.1093/isd/ixab017 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE240C-FFC7-FF8C-FC8B-B79165C6E47F |
|
treatment provided by |
Felipe |
|
scientific name |
Chazara briseis |
| status |
|
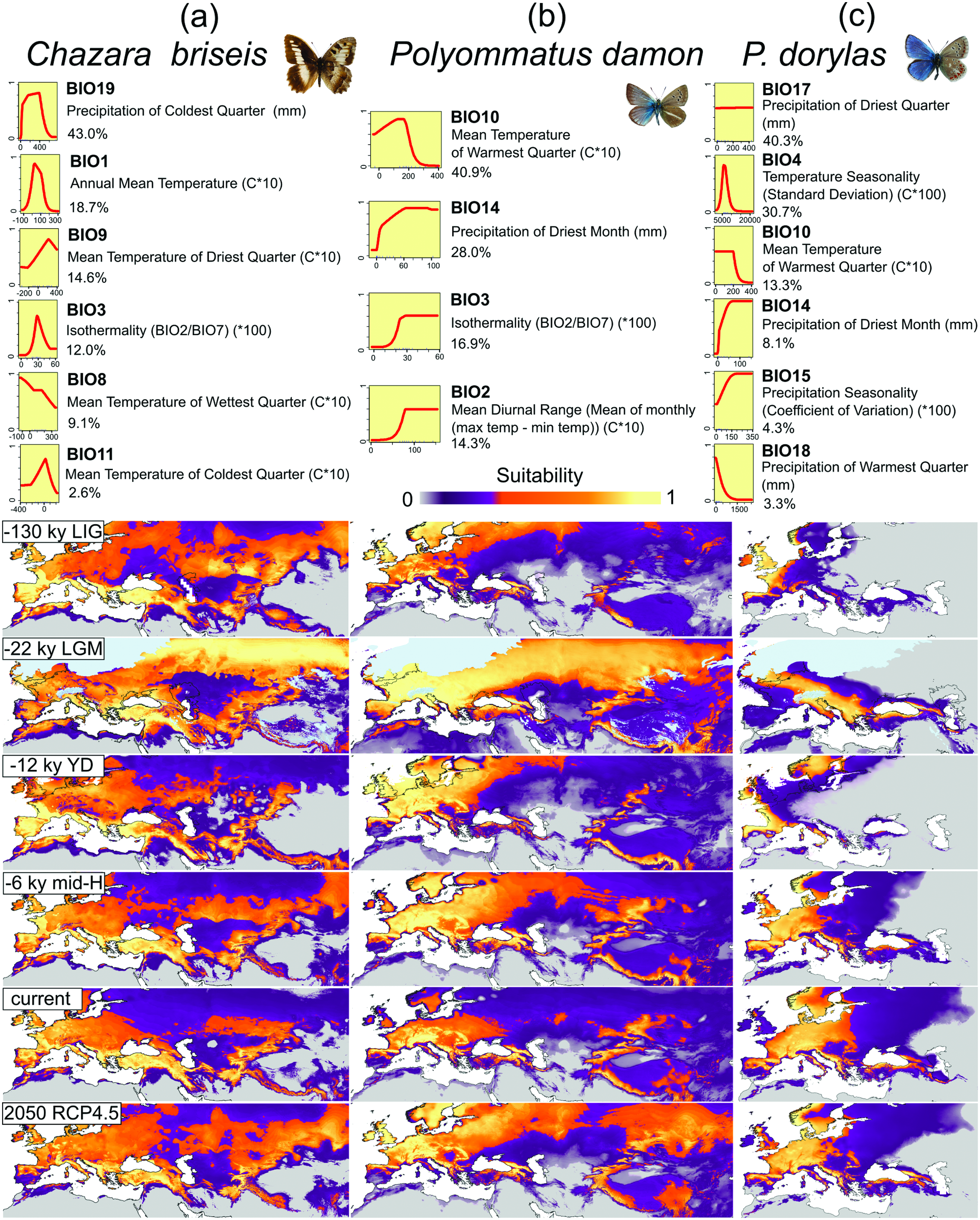
Chazara briseis Biogeography
The populations in Central Europe belong to a mitochondrial group (BAPSb1) widely distributed from Spain to the Altai region, distinctly separated from the populations in North Africa, and the samples from south-eastern parts of the range. The widely distributed group and south-eastern group display evidence of secondary contact ( Fig. 2 View Fig ). The BAPSb1 consists of a haplotype B1 present through the entire region, and unique haplotypes at almost all sites sampled. The observed population expansion could be a possible response to an expansion of steppe grasslands during the Pleistocene. Such structure (a common haplotype with many singleton forms), however, is a frequently observed phenomenon ( Dincă et al. 2021), implying an impossibility to sample all the unique forms, and might be a result of cyclic population contractions and expansions.
The local populations in Central Europe are relatively distinctive, suggesting some level of relatively long-term isolation: populations from DE (haplotypes B4 and B7) are related to those from Western Europe; CZ populations harbor one haplotype unique for the region (B19) and a haplotype shared with HU and AT (B20). Populations from AT further consisted of the widespread B1 and a unique haplotype B22.
The more recent past was investigated by allozyme studies by Johannesen et al. (1997) in DE (Halle region, Saxony-Anhalt) and
Kadlec et al. (2010) in CZ. Both studies targeted metapopulations restricted to a few remnant sites, the latter containing ≈2,000 adults. Both disclosed high within-population diversity, comparable with some widely distributed species. This was interpreted as a sign of recent contractions of formerly abundant and connected populations, not yet reflected in the studied marker.
Climatic models ( Fig. 6 View Fig ) revealed that a climate appropriate for C. briseis was widespread across the Western Palearctic, including Central Europe, for upper Pleistocene glacial cycles, with retreats from mountains and deserts during glacial maxima. In the Holocene and the last interglacial, the climatic niche became fragmented by avoidance of northeastern lowlands, with the most suitable areas existing in Western and Southern Europe, Anatolia and Central Asia. Hence, the expansion observed in the molecular population structure is likely of pre-Holocene origin, although we cannot decide whether this happened during the last glacial or earlier. C. briseis is thus climatically linked with both cold glacial steppe and open habitats of more southerly regions. This requires a high climatic tolerance, documented in experiments with larval overwintering (Vrba et al., unpublished data). The larvae survived low temperatures comparable with alpine butterflies (winter supercooling point: –17°C) and employed a mixed cold hardiness strategy, with some larvae surviving and some avoiding body fluids freezing.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.