
Echiniscus meridionalis, Gąsiorek, 2023
|
publication ID |
https://doi.org/ 10.1016/j.jcz.2023.02.004 |
|
publication LSID |
lsid:zoobank.org:pub:8A36A0BF-AA4F-427F-980B-E70DE960695E |
|
DOI |
https://doi.org/10.5281/zenodo.8171597 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE752F-FF94-FFD8-FCA0-F950FE1BFAF4 |
|
treatment provided by |
Felipe |
|
scientific name |
Echiniscus meridionalis |
| status |
stat. nov. |
3.2. Redescription of Echiniscus meridionalis View in CoL stat. rev.
Figs. 2 View Fig – 7 View Fig View Fig View Fig , Tables 2–4 View Table 2 View Table 3 , Supplementary Material 4 with raw morphometric data.
ZooBank registration number: urn:lsid:zoobank.org:pub:8A36A0BF- AA4F–427F–980B-E70DE960695E.
3.2.1. Terra typica
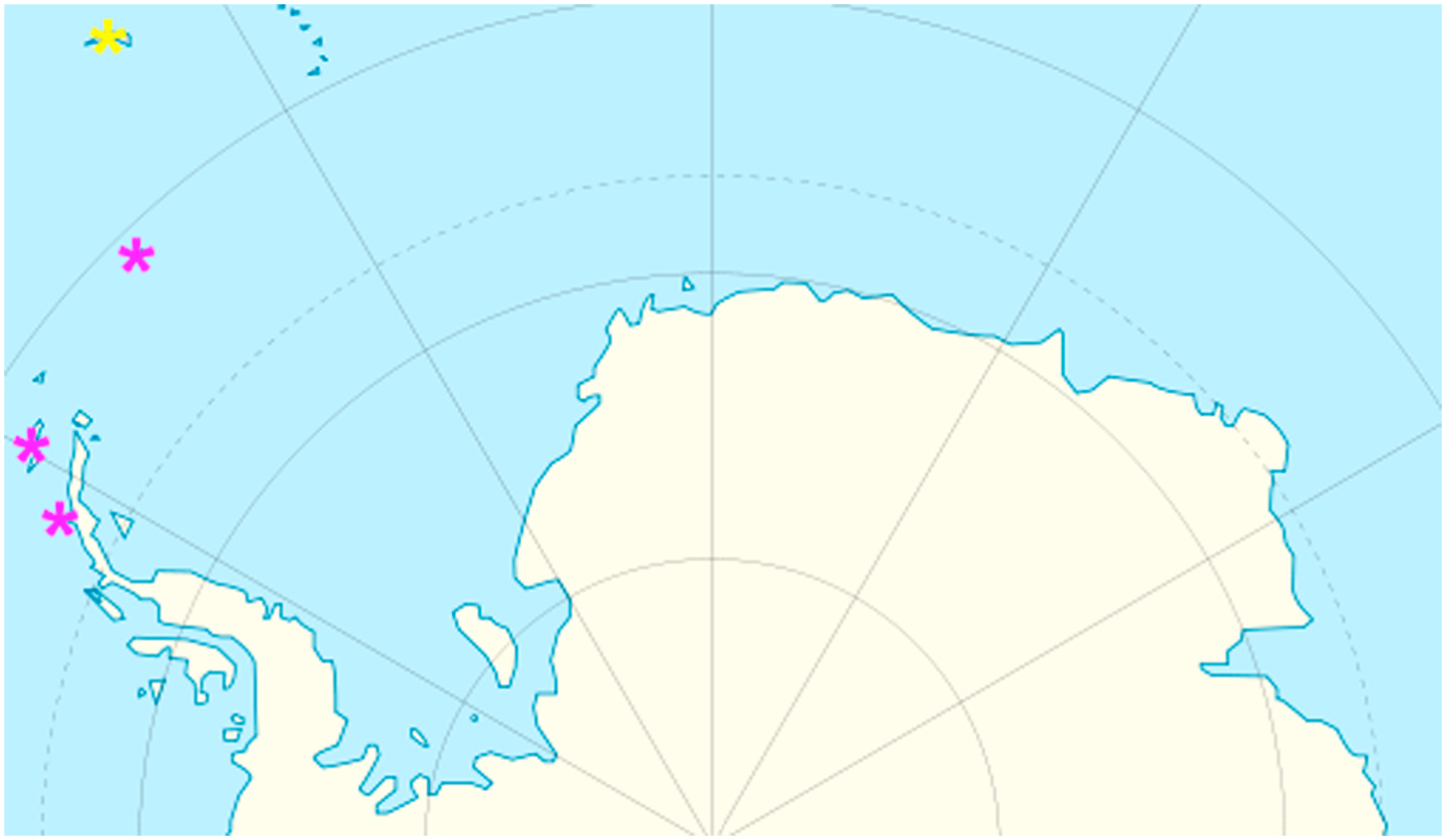
South Orkney Islands. Material examined. South Shetland Islands (see Table 1 View Table 1 for detailed data).
3.2.2. Etymology
From Latin meridionalis = southern, referring to the distribution of the species in Antarctic and sub-Antarctic archipelagos. An adjective in nominative singular.
3.2.3. Adult females (i.e. from the third instar onwards; measurements and statistics in Table 2 View Table 2 )
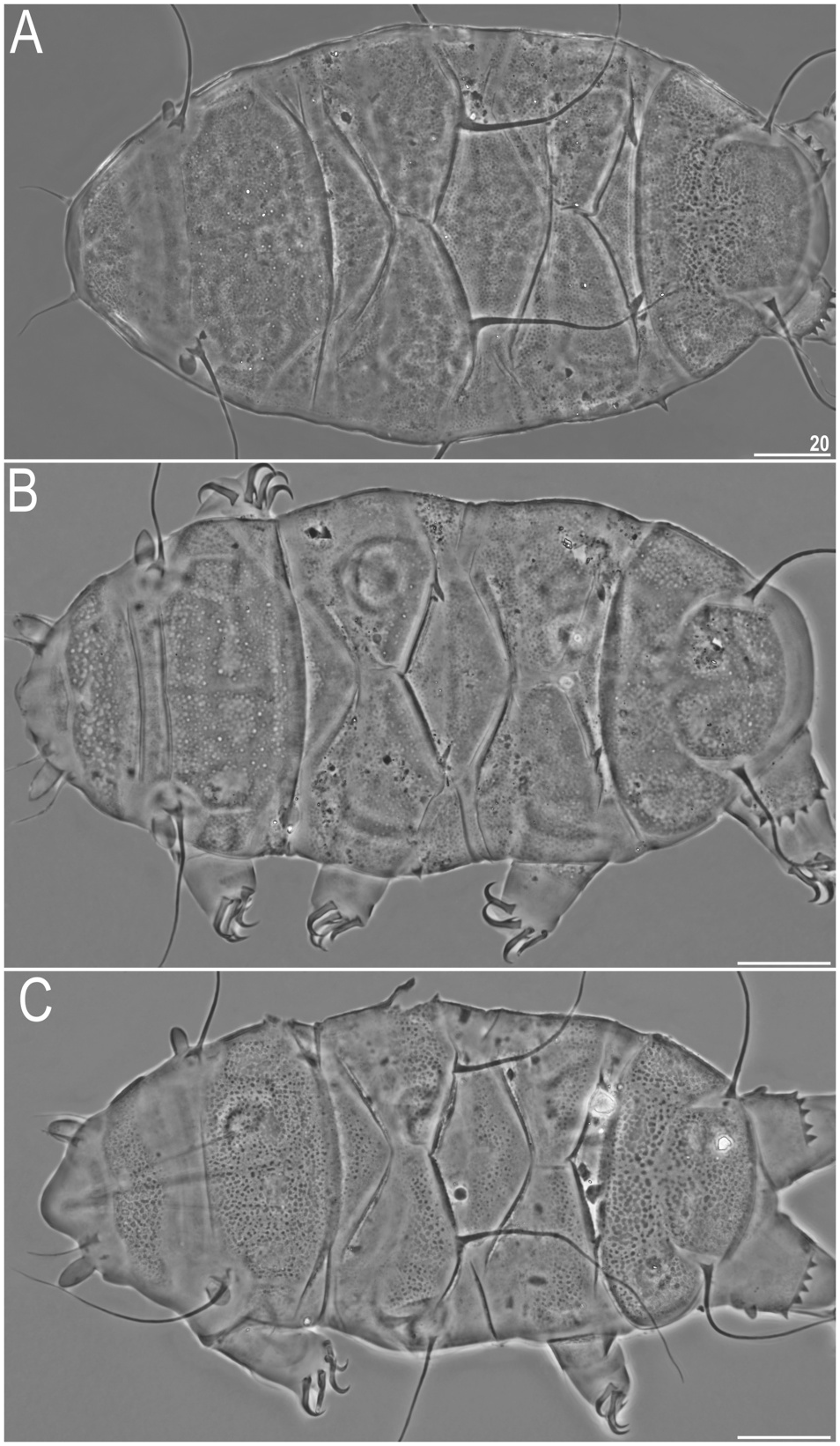
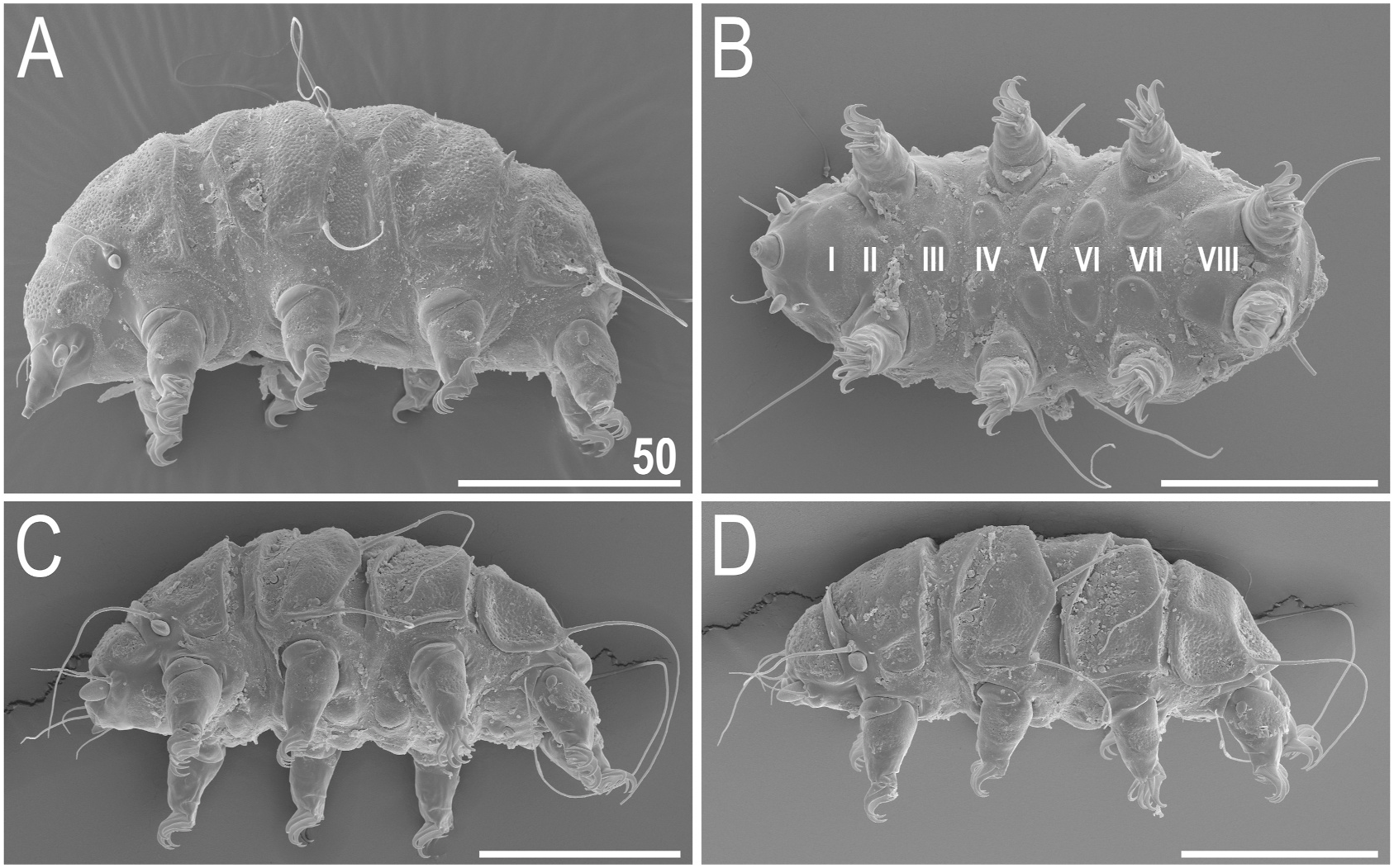
Body small and plump ( Figs. 2A View Fig and 3A View Fig ), light yellow to dark orange in alive specimens. Eyes dark red, minute and dissolving immediately in Hoyer’ s medium. Peribuccal cirri relatively long with respect to the body length ( Figs. 2A View Fig , 3A View Fig and 4A View Fig ); cirrus A of a moderate length, i.e. reaching approximately 1/3 of the body length; all cephalic cirri with well-delineated cirrophores. Cephalic papillae (secondary clavae) slightly longer than (primary) clavae, both barrel-shaped ( Fig. 4A View Fig ). Body appendage configuration A-C-C d -D-D d -E, with most appendages formed as long, flexible cirri. Short, but robust spines present in positions D and D d ( Figs. 2A View Fig and 3A View Fig ).
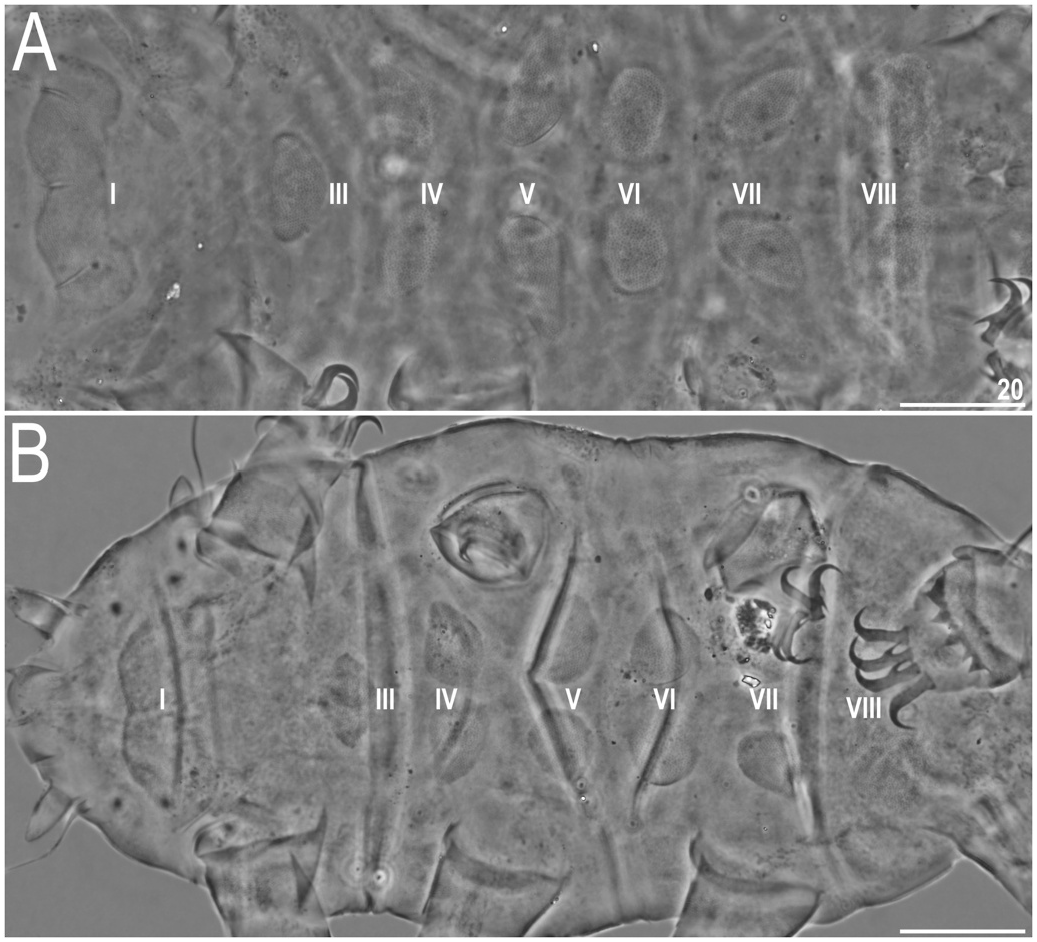
Dorsal plate sculpturing comprises: (I) poorly elevated epicuticular granules, with a strong tendency to merging in all plate portions but the anterior portions of paired segmental plates I–II, median plate 2, and the much reduced median plate 3, where they are separate ( Figs. 2A View Fig and 3A View Fig ), ( II) pseudopores in the form of delicate depressions distributed irregularly throughout the dorsum, and ( III) pores, present mainly in the cephalic, scapular ( Figs. 2A View Fig and 3A View Fig ), and occasionally in the caudal (terminal) plates. Pores may be absent as well. Cephalic plate narrow, with the usual chalice-like anterior incision. Cervical plate even narrower, but clearly present and rectangular, tightly joined with the scapular plate. Scapular plate with a distinct faceting-like pattern formed by a system of poorly elevated ridges ( Figs. 2A View Fig , 3A View Fig and 4A View Fig ); lateralmost plate portions weakly demarcated from the central portion by a similar ridge ( Fig. 4A View Fig ) that replaced a usual suture/incision. Median plate 1 unipartite, m2 bipartite and much larger, m3 developed as a triangular sculptured surface dominated by the caudal plate. Paired segmental plates I–II identical in size and shape. Caudal plate with large triangular posterior incisions ( Fig. 2A View Fig ) that are joined by a central semicircular ridge ( Figs. 2A View Fig , 3A View Fig and 4A View Fig ). A weakly marked ridge perpendicular to the former may be developed, delineating a cross-like epicuticular structure.
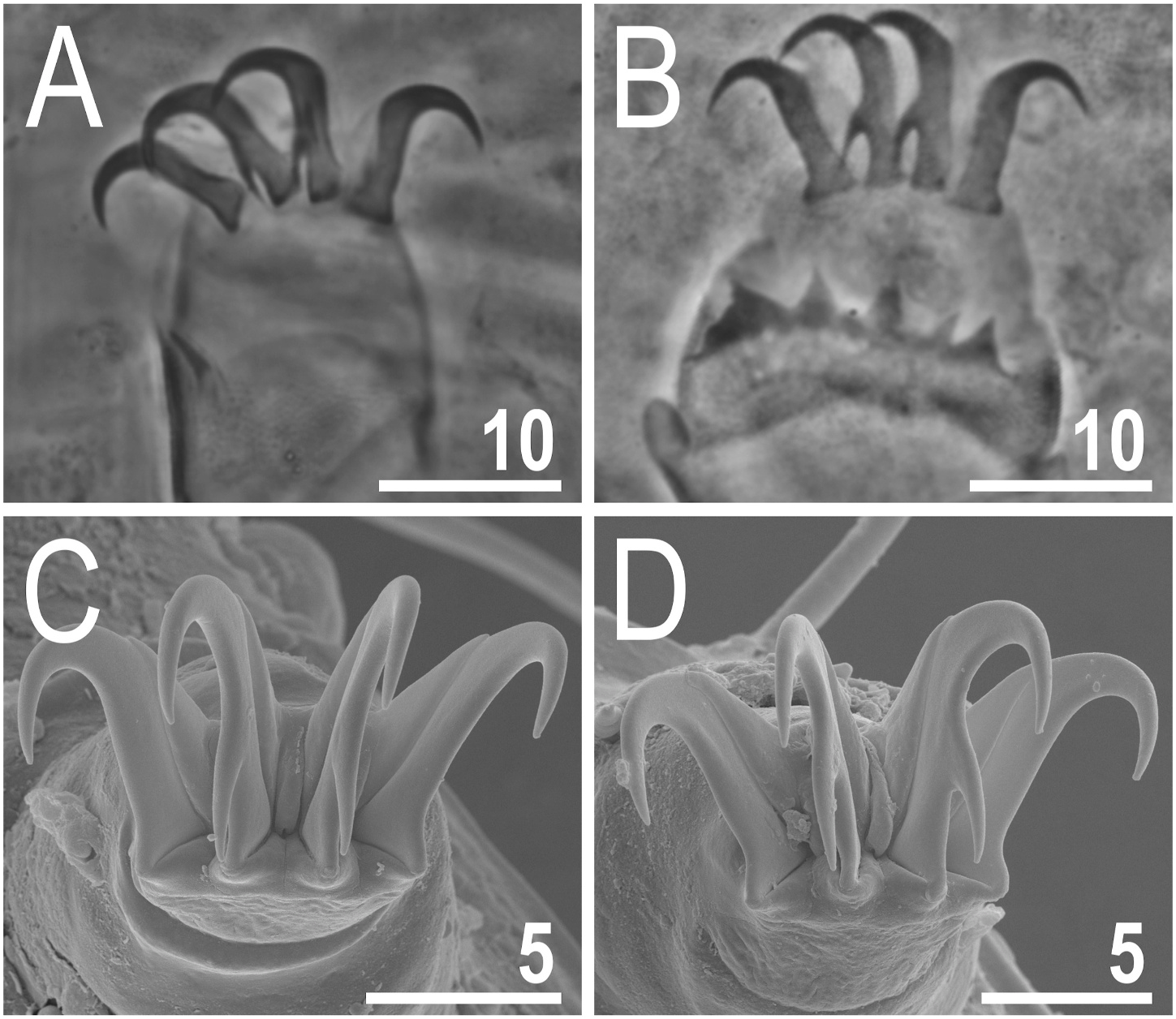
Ventral plate armature variously developed, as plates in some individuals may be almost indiscernible, but the general plate formula stands as VIII:2-1-1-2-2-2-2-2 ( Figs. 4B View Fig and 5A View Fig ). Particularly the plate in the second row can be hardly identifiable in PCM ( Fig. 5A View Fig ). Ventral plates are smooth ( Fig. 4B View Fig ), but the endocuticular pillars are easily observable in PCM ( Fig. 5A View Fig ). Besides, no granulation present on venter. Hexapartite gonopore and tripartite, large anus ( Figs. 4B View Fig and 5B View Fig ). Legs with evident pulvini embedded in proximal portions of limbs and pedal plates in median portions, developed as wrinkled areas with sparse and occasional pores ( Figs. 4A View Fig and 6B View Fig ). Long spine I present ( Figs. 4A View Fig and 6A View Fig ); papilla IV large and barrel-shaped, dentate fringe on leg IV with numerous irregular teeth, typically interconnected at their bases ( Figs. 2A View Fig , 4A View Fig and 6B View Fig ). Claws with branches typically curved, those of legs IV slightly higher than the remaining ones. Claw spurs heteromorphic, as spurs IV are more divergent from the branches compared with spurs I– III. The size and position of spurs on claw branches (ca. 1/3–2/5 of the claw height) identical on all claws ( Fig. 6 View Fig ).
3.2.4. Adult males (i.e. from the second/third instar onwards, see remarks; measurements and statistics in Table 3 View Table 3 )
Sexual dimorphism evinces in several characters of males: thinner and shorter body ( Figs. 2B and C View Fig , 3B View Fig , 4C–D View Fig ), prominent and swollen clavae ( Figs. 2B and C View Fig , 3B View Fig , 4C–D View Fig and 5B View Fig ), and often better visible (darker in PCM) ventral plates ( Fig. 5B View Fig ). Rarely, all appendages but A and E may be formed as spines ( Fig. 2B View Fig ). Gonopore circular.
3.2.5. Juveniles (i.e. the second instar; measurements and statistics in Table 4)
Qualitatively similar to sexually mature females, and smaller than both sexes. Gonopore absent.
3.2.6. Larvae (i.e. the first instar; measurements and statistics in Table 4)
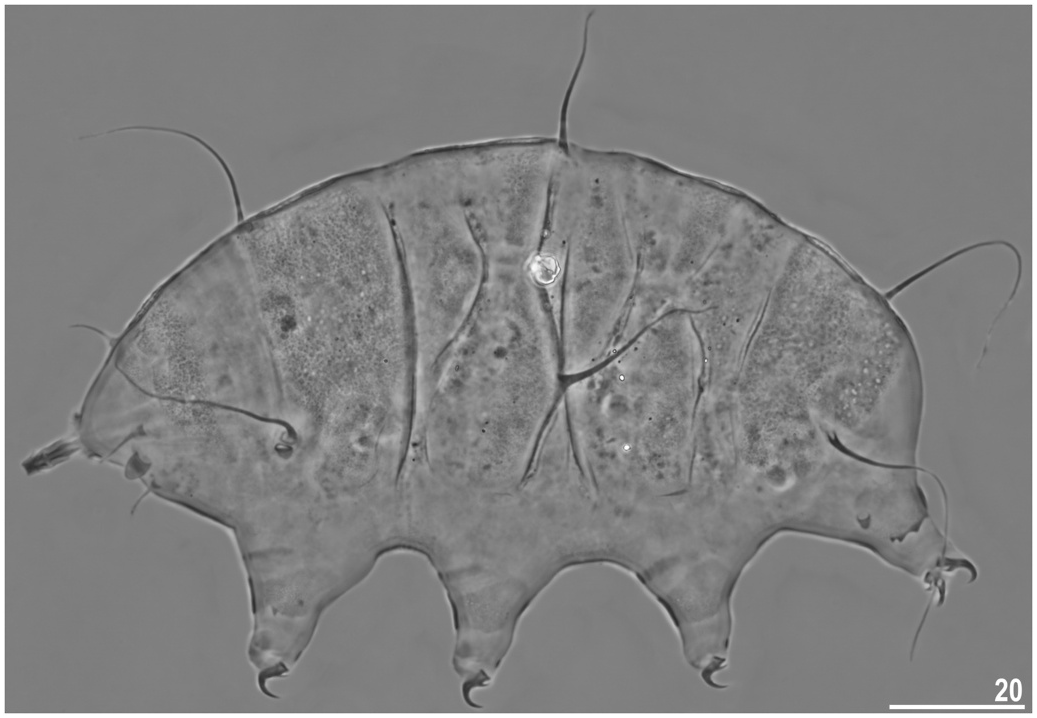
Qualitatively similar to later life stages, but smaller than juveniles. Body appendage formula: A-Cd- E, all formed as cirri ( Fig. 7 View Fig ). Two claws per leg. Gonopore and anus absent.
3.2.7. DNA markers
Single haplotype was found in 18S rRNA ( OQ 472387), 28S rRNA ( OQ 472389), ITS-1 ( OQ 476878), ITS-2 ( OQ 476879), and two in COI ( OQ 472391–2; p -distance = 0.2%).
3.2.8. Differential diagnosis
A strong sexual dimorphism, the unusual dorsal plate sculpturing (in some specimens, epicuticular granules may be barely elevated above the surface of plates, and in an initial observation in PCM such exemplars seem to be sculptureless), and the ventral armature covering an entire venter, replacing the endocuticular granulation typical for Echiniscus , together with the geographic range embracing solely Antarctic and sub-Antarctic islands ( Fig. 8 View Fig ), make E. meridionalis stat. rev. easily recognisable and well-differentiated morphologically from all other Echiniscus spp.
3.2.9. Remarks
The species can be frequently encountered on King George Island, being one of the two commonest echiniscids in that location (the other is Barbaria jenningsi , often co-inhabiting the same mosses and lichens, see Table 1 View Table 1 ). An undescribed Pseudechiniscus species, eutardigrades Acutuncus antarcticus , Diphascon rudnickii , and unidentified species of the Mesobiotus furciger group ( Short et al., 2022) are other relatively common taxa that co-occur with E. meridionalis stat. rev. However, irrespective of its ubiquity in the area, the population density must be low given that typically few representatives were found in samples ( Table 1 View Table 1 ). Ventral armature is already developed in E. meridionalis stat. rev. at sexually immature life stages (i.e. in larvae and juveniles), which often does not take place in the Echiniscidae .

| PCM |
Polish Collection of Microorganisms |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |