
Paraxenylla sooretamensis, Queiroz, Gabriel C. & Deharveng, Louis, 2008
|
publication ID |
https://doi.org/10.5281/zenodo.183626 |
|
DOI |
https://doi.org/10.5281/zenodo.5615627 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE87B8-FFBB-8E1A-01F8-FA4AAF47FAFE |
|
treatment provided by |
Plazi |
|
scientific name |
Paraxenylla sooretamensis |
| status |
sp. nov. |
Paraxenylla sooretamensis sp. nov.
Figs 1–9 View FIGURES 1 – 9 , Tabs 1–3 View TABLE 1 View TABLE 2 View TABLE 3
Type locality. Sooretama, Espírito Santo, Brazil; 40 km from coast. Forest litter of Atlantic Forest.
Type material. 10 April 2007 Gabriel Queiroz leg. Holotype female and 6 paratypes (Nº 1829 CM/ MNRJ) on slide at Museu Nacional do Rio de Janeiro, Brasil.
Etymology. From the name of the type locality: Sooretama, a word of the Indian language Tupi that means “home of the wild animals”.
Description. Body length ( 7 specimens examined): 0.60 mm ( holotype) and 0.44–0.82 mm ( paratypes). Color dark blue. Dorsal chaetotaxy of short subequal mesochaetae and long S-chaetae.
Ratio head: antenna = 0.64. Ant I with 7 chaetae. Ant II with 11 chaetae. Sensory organ of Ant III with 5 S-chaetae: 2 small internal rods S3 and S4 behind a fold of the integument, 2 longer guard chaetae S2 and S5, and one external microchaeta S1. Ant IV with 4 thick S-chaetae and a small S-microchaeta between the 3 Schaetae of the external group. Subapical organ present as a very short burried chaeta continued into the integument ( Fig. 1 View FIGURES 1 – 9 ). Apical bulb simple.
5+5 eyes pigmented. Labral formula: 24/554 chaetae. Labial palp chaetotaxy strongly reduced, with (tentative homologies after Fjellberg 1999): A and B as straight, thin papillated chaetae; C, D and E not developed; 10 guard chaetae, including 4 large bent flattened chaetae arranged as a distal file (b4, d4, d3 and e3), 2 medium size chaetae of uncertain homology and 4 microchaetae (a1, b2, d2 and one of uncertain homology); 6 proximal chaetae; and 3 hypostomal chaetae (H, h1 and h2) ( Fig. 2 View FIGURES 1 – 9 ). Mandible head thin, its distal part subparallel, with 4 subequal apical teeth ( Fig. 3 View FIGURES 1 – 9 ). Maxillary capitulum ( Fig. 4 View FIGURES 1 – 9 ) with 5 fringed lamellae: 3 of medium-size, one short and one hypertrophied; a sixth internal lamella appears smooth and may actually correspond to the ungulum. Maxillary outer lobe reduced, without sublobal hairs ( Fig. 5 View FIGURES 1 – 9 ).
Thoracic tergum I with 3+3 chaetae. No clearly differentiated macrochaetae on tergites. Dorsal chaetotaxy of head, thorax and abdomen as in Fig. 6 View FIGURES 1 – 9 . Dorso-internal S-chaetae in position 33/5555. Axial chaetae 66/ 4446. Chaetae m1 present, m2 absent on Th II–III; p2 on Abd I–III and a1 on Abd V as other ordinary chaetae.
Unguis with a small inner tooth on legs I, II and III. Two minute median lateral teeth seems to be present, only visible when the claw is in favourable position. Tibiotarsi without clavate tenent hairs ( Fig. 8 View FIGURES 1 – 9 ). Leg chaetotaxy summarized in Tab. 2 View TABLE 2 .
Linea ventralis distinct on thoracic sternites, reaching the ventral tube. Ventral tube with 4+4 chaetae. Tenaculum with 3 teeth on each ramus. Mucrodens well developed ( Fig. 7 View FIGURES 1 – 9 ). Manubrium with a bilobed ventro-distal sclerification on each side corresponding to the articulations dens-manubrium. Dens long, with two ventral chaetae and, dorsally, a large longitudinal strip devoid of primary granules; mucro thin and long, tapering, separate from the dens and without lamella. Dens 2–2.5 times longer than mucro. Abd VI dorsally with 5+5 chaetae, one uneven posterior chaeta, and 2 microchaetae An on the supra-anal valve ( Fig. 9 View FIGURES 1 – 9 a); ventrally, with 13+13 chaetae Ve, one uneven posterior chaeta Ve, and a single microchaeta An on each lateral valve, located near the axis ( Fig. 9 View FIGURES 1 – 9 b). Anus terminal, no longitudinal furrow between the lateral anal valves. No anal spines.
Discussion. All species of the genus Paraxenylla known so far were found in marine littoral habitats ( Palacios-Vargas & Janssens 2006), including P. cubana (Palacios-Vargas comm. pers.). Surprisingly, P. sooretamensis sp. nov. was found in the forest litter of the Table land Atlantic Forest of southeast Brazil, 40 km away from the coast line.
The new species is well characterized in the genus ( Tab. 3 View TABLE 3 ). Its mouthparts can only be compared with those of P. affiniformis for which detailed descriptions are available ( Gama & Deharveng 1984, Jordana et al. 1997, Fjellberg 1999). Both species have many peculiar characters in common: thin mandible, hypertrophied lamellae of the maxilla, labial palp chaetotaxy very reduced. However, a closer examination shows that the modifications of the maxilla is different in the two species (in particular, most of the fringed lamellae are much larger in P. affiniformis ). In addition, labial palp chaetae, though difficult to homologize with the patterns provided by Fjellberg (1999), are arranged very differently in P. affiniformis compared to P. sooretamensis , with, among other differences, more large distal chaetae (5–6 against four) and less microchaetae (two against four) in P.affiniformis . Mouthparts of two other species ( P. a re n o s a and P. mangle ) are also reduced in a similar way, but have been described too succinctly to allow comparison. P. mangle differs from P. sooretamensis by a shorter mucro, chaeta S2 of Ant III organite shift between S3 and S4, m1 absent on Th II–III and 2+2 chaetae on the ventral tube. P. arenosa differs from P. sooretamensis by the widening of some maxillary lamellae, a trilobed apical bulb, a longer mucro and 3+3 chaetae on the ventral tube.
The guard chaetae of the Ant III organite of the new species are external to the two small rods as in most Hypogastruridae , while in P. affiniformis , P. mangle , P. peruensis and P. p i l o u a one of the guard chaetae is located between the rods.
Linea ventralis is not visible in P. affiniformis according to Gama and Deharveng (1984), contrary to our new species. It is not described in other species.
P. oceanica and the new species are the only ones in the genus to have 4+4 chaetae on ventral tube, which is the usual number in Hypogastruridae . Other Paraxenylla have 3+3 to 1+1. P. oceanica differs from P. sooretamensis only by its mucro fused to the dens, but most chaetotaxic and mouthpart characters remain undescribed in this species, which has to be considered as species inquirenda.
By its dorsal chaetotaxy, resembling that of Xenylla in many respects ( Tab. 1 View TABLE 1 ), Paraxenylla sooretamensis sp. nov. is close to P. arenosa ( Gama 1980, 1981) and totally apart from other species of the genus. The placement of these two species in the genus Paraxenylla will have to be re-examined when more information will be available on the mouthparts and chaetotaxy of other species of the genus. They represent in any case a well defined taxonomic group intermediate in several respects between Xenylla (by their chaetotaxy) and Paraxenylla (by their mouthparts, absence of anal spines and anus in a terminal position).
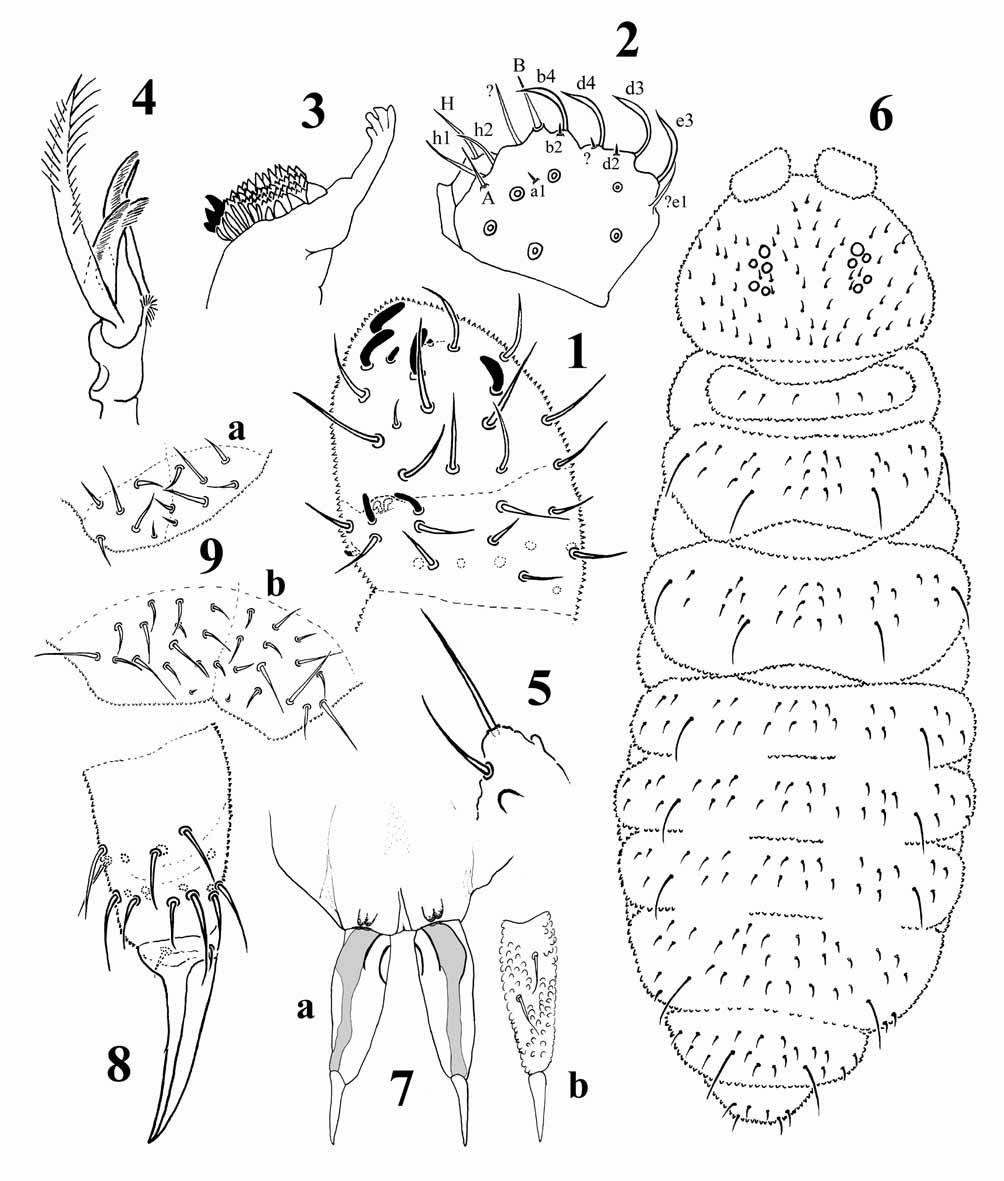
TABLE 2. Chaetotaxy of legs.
| Tita | Fe | Tr | Cx | Scx2 | Scx1 | |
|---|---|---|---|---|---|---|
| Leg I | 19 | 12 | 5 | 3 | 0 | 1 |
| Leg II | 19 | 12 | 5 | 7 | 2 | 2 |
| Leg III | 18 | 10 | 4 | 8 | 2 | 3 |
TABLE 3. Some morphological characters of the species of Paraxenylla (in part from Palacios-Vargas & Janssens 2006) (na: not applicable).
| Axial chaetae | m1 on Abd IV | VT chaetae | Tenac. teeth | Tenent hairs on Tita III | Unguis Tooth | |
|---|---|---|---|---|---|---|
| affiniformis | 44/44464 | near a1 | 1+1 | 3+3 | 5 | + |
| arenosa | 66/44464 | between a1 and p1 | 3+3 | 4+4 | 1 | + |
| cubana | 44/?4442 | na | 1+1 | 2+2 | - | - |
| lapazana | 44/44442 | na | 1+1 | 3+3 | 3 | - |
| mangle | 44/44462 | between a1 and p1 | 2+2 | 3+3 | 3 | - |
| oceanica | ? | ? | 4+4 | 3+3 | 3 | - |
| peruensis | 44/44462 | near a1 | 1+1 | 3+3 | 4 | - |
| piloua | 44/44442 | na | 1+1 | 3+3 | 4 | - |
| sooretamensis sp. nov. | 66/44464 | between a1 and p1 | 4+4 | 3+3 | - | + |
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |