
Atopostroma Yang & Dong, 1979
|
publication ID |
https://doi.org/10.3853/j.0067-1975.60.2008.1497 |
|
persistent identifier |
https://treatment.plazi.org/id/03EF4E2E-8076-9D1C-93E7-F749FB2BF831 |
|
treatment provided by |
Carolina |
|
scientific name |
Atopostroma Yang & Dong, 1979 |
| status |
|
Type species. Atopostroma tuntouense Yang & Dong, 1979 .
Diagnosis: Pachysteles (pillars, in poorly preserved condition) moderately thick, superposed, inverted conical to post- and spool-shaped; microstructure usually microreticular. Pachystromes (seem to be laminae in less well preserved state) are laterally extensive and variably thickened; differentiated into thin, sharply defined, flattened, laterally persistent, microcolliculate bounding microlamina at top and, where well enough preserved, underlain by irregularly spread network of microreticular material that joins pachysteles, and may include other close-spaced, but more discontinuous, microlaminae; pachystrome floors irregular, best defined in dome-shaped contacts over galleries. Dissepiments rare, widely scattered, either small cyst-like, or long, wavy, microlamina-like plates. Microreticular structure of skeleton composed of variable fine network of micropillars and microcolliculi in microlaminae; in tangential section microstructures composed of acosmoreticular material; microreticular features of pachysteles and pachystromes are also largely acosmoreticular in longitudinal section, except clinoreticular where pachystele margins diverge upwards; sometimes microreticular features masked by diagenetic alteration, leaving skeletal elements preserved predominantly with melanospheric to compact microstructures.
Discussion. Yang & Dong’s (1979, p. 74) original generic diagnosis included: laminae thin, compact, without thickened material; pillars thick, upward spreading, superposed across a few laminae, with connecting radial processes; astrorhizae rudimentary; no “pseudozooidal” tubes [=allotubes]; microstructure cellular or maculate. The maculae were apparently darker spheres on a lighter background (equivalent to the melanospheres of Stearn, 1965, 1966). Though resembling Actinostroma (given the recognition of hexactinellid type “connecting processes” of actinostromatids), Yang & Dong preferred to group Atopostroma as belonging to family Stromatoporidae , and this position was maintained by Dong & Wang (1982) and Wang (1988) in subsequent descriptions of Chinese species. Stearn (1980, 1983) on the other hand favoured a grouping of Atopostroma within the Clathrodictyidae because of the characteristic spread of thick, superposed, pillars beneath successive, thin, single-layered laminae (or microlaminae), which suggested a close relationship with clathrodictyid genus Gerronostroma . Also, Stearn (1966, p. 101) suggested Actinostroma stellulatum distans Ripper, 1937 as a species of Gerronostroma (see further discussion below). Mistiaen (1985) however argued that the microstructure in Gerronostroma was not the same, and preferred a separate grouping for Atopostroma .
Later, Stearn (1990) identified a clinoreticulate microstructure in “pillars” of a form he called Atopostroma tuntouense from Stuart Bay, Bathurst Island, in Arctic Canada, but not identical with the type species—see Webby et al. (1993, p. 173). Additionally, Stearn (1993) reassigned Atopostroma to the family Syringostromatidae based on the presence of a microreticulate microstructure within the order Syringostromatida . Webby & Zhen (1993) and Webby et al. (1993) then followed Stearn’s lead, reinterpreting the taxon Actinostroma stellulatum distans Ripper, 1937 , as another valid representative of genus Atopostroma . Others—see Prosh & Stearn (1996), Stearn, in Stearn et al. (1999), and May (1999, 2005)—have maintained the grouping of Atopostroma within the family Syringostromatidae (order Syringostromatida ).As mentioned earlier, Stearn (1993) also commented on the two genera exhibiting transitional forms in the late Lochkovian Martins Well Limestone Member of north Queensland.
Dong (2001, p. 150), after presenting a similar generic diagnosis of Atopostroma to the original diagnosis of Yang & Dong (1979, p. 74), adopted a different approach to classifying the genus. Instead of the earlier Yang & Dong assignment to the “stromatoporids”, Dong (2001), perhaps, after giving greater weight to the presence of long pillars and “connecting rods” forming hexactinellid networks, placed Atopostroma in the order Actinostromatida , though his conception of the order remained markedly different from groupings adopted by others (see, for example, Stock, in Stearn et al., 1999). Dong (2001) included the genus Atopostroma in the family Gerronostromatidae Bogoyavlenskaya, 1969 along with five other genera, and he also transferred that family to the order Actinostromatida . The genera he included altogether in the family were of mixed affinities—clathrodictyid ( Gerronostroma, Pseudoactinodictyon, Clathrostroma ), stromatoporellid ( Amnestostroma ), syringostromatid ( Atopostroma ) and problematical ( Pseudostromatopora )—and none of them, with the possible exception of Atopostroma , displays features of the Actinostromatida (see Stock, in Stearn et al., 1999 p. 32). Only Atopostroma with its microreticulate microstructure could possibly be interpreted as an actinostromatid (but here it is preferentially treated as a part of the order Syringostromatida ). Only approximately half of Dong’s (2001) listed genera belong to the family Gerronostromatidae , and that family, according to Nestor ( in Stearn et al., 1999, p. 23), belongs to the order Clathrodictyida . All the families of the Clathrodictyida developed a compact skeletal microstructure.
The genus Atopostroma has proven to exhibit an original microreticular microstructure, and it only apparently produces a compact or melanospheric (= Dong’s “maculae”) microstructure when affected by secondary diagenetic processes. Here, we treat the primary skeletal microreticulation as having a primary focus in the classification of genus Atopostroma , and therefore the genus is as an indubitable member of the order Syringostromatida . It may have a link to actinostromatids given that some taxa exhibit small-scale structures such as micropillars and microcolliculi, like the genus Densastroma Flügel, 1959 (e.g., within the family Densastromatidae Bogoyavlenskaya, 1974 ), but it is not closely related to a genus like Actinostroma that has long pillars with a compact microstructure, as suggested by Dong (2001).
Species of Atopostroma are widely distributed in Devonian sequences worldwide: in particular they are reported from the Lower Devonian (Lochkovian) of Arctic Canada and New York, USA ( Stearn, 1990; Stock, 1997); in the Lower Devonian (Pragian) of Bohemia, Czech Republic ( Pocta, 1894; May, 1999, 2005); in the Lower Devonian (Pragian-Emsian) of eastern Australia ( Ripper, 1937; Webby & Zhen, 1993, Webby et al., 1993; and herein); Lower Devonian (Emsian) of Arctic Canada and Yukon ( Stearn, 1983; Prosh & Stearn, 1996), and from Guangxi and Sichuan in China (Yang & Dong, 1979; Dong, 2001); as well as from the Middle Devonian of the Kuznetsk Basin, Russia ( Yavorsky, 1955), Yunnan, in China ( Dong, 2001) and possibly from Afghanistan ( Mistiaen, 1985).
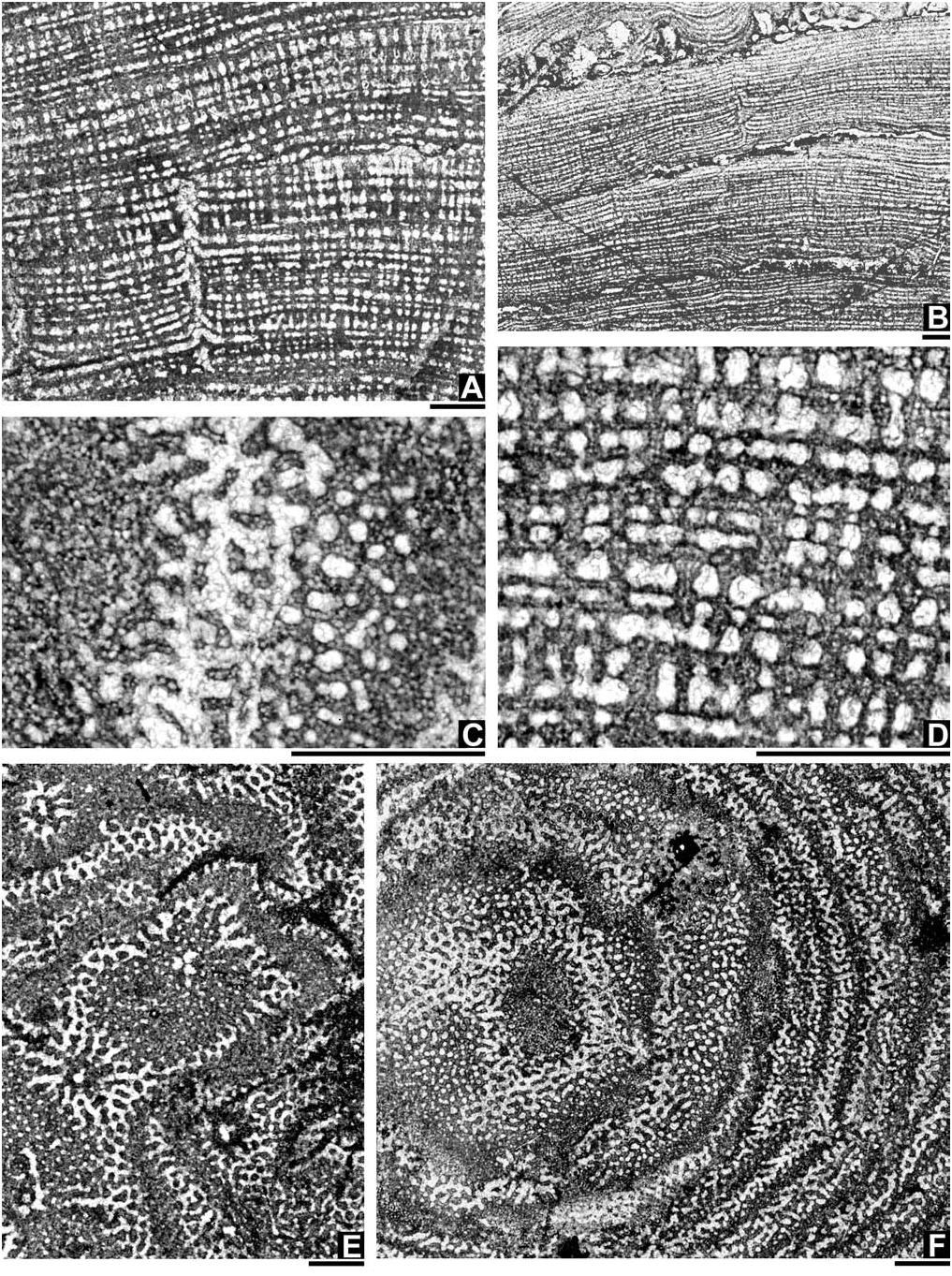
[ Fig. 3 View Fig continued] … gallery space that grades up into an area defined by a mix of interconnected microreticulate material between small, enclosed spar-filled galleries, and darker, denser, pachystromal layers with a more uniform microreticulation of differentiated micropillars that interconnect to microcolliculi around tiny microgallery spaces towards top, x35; ( D) longitudinal section, AM F.134879 (AM FT.15066), showing a acosmoreticular to clinoreticular microstructure, upwardly expanded to hour-glass shaped, superposed pachysteles, and a somewhat irregularly thickened pachystromal layer of microlaminae with the uppermost microlamina usually having the greatest continuity, ×35; ( E) tangential section, AM F.134882 (AM FT.15073) from the Lomandra Limestone of SD170 section, 274m above the base, ×10; F, tangential section, AM F.134879 (AM FT.15067), ×10—for detail see Fig. 3C View Fig . All scale bars = 1 mm.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.