
Campylothorax sabanus
|
publication ID |
https://doi.org/10.1080/00222933.2016.1145272 |
|
publication LSID |
urn:lsid:zoobank.org:pub:07C2E4C4-7C51-48C7-8D45-7E6B9442C497 |
|
DOI |
https://doi.org/10.5281/zenodo.5628361 |
|
persistent identifier |
https://treatment.plazi.org/id/03F17166-D01A-EE0A-FF15-FD05FBE7FB8E |
|
treatment provided by |
Plazi |
|
scientific name |
Campylothorax sabanus |
| status |
|
First-instar tergal chaetotaxy of Campylothorax sabanus
First-instar and adult C. sabanus described below were collected in Puerto Rico, Isabela, Guajataca Commonwealth Forest, leaf litter on summit of mogote at intersection of trails 25, 26 and 27, 3 January 2014, F. Soto and H. Soto, two first instar and two adults. Additional observations on adult C. sabanus morphology are based on material collected at Puerto Rico, Mayagüez, Cerro Las Mesas, leaf litter along road, 23 October 1997, F. Soto, one specimen; Maricao, Monte del Estado Commonwealth Forest, leaf litter near observation tower on Rd. 120, 8 June 1998, F. Soto, two specimens; Utuado, Rio Abajo Commonwealth Forest, leaf litter on summit of mogote 10 January 2014, F. Soto, two specimens. US Virgin Islands, St. Thomas, patch of forest along Rd. 40, 20 June 2000, F. Soto, one specimen; St. John, Cinnamon trail, 30 June 2000, F. Soto, two specimens.
Determination of primary chaetotaxy homology for C. sabanus is based on comparisons with the chaetotaxy reported in the literature for Trogolaphysa jataca ( Wray, 1953) (Soto- Adames 2015), Pseudosinella decipiens Denis, 1924 and P. impedens Gisin & Gama, 1969 ( Barra 1975) , Seira dowlingi ( Wray, 1953) ( Soto-Adames 2008) , Homidia jordanai Pan, Shi and Zhang 2011 , Orchesella fl avescens ( Bourlet, 1839), Heteromurus nitidus ( Templeton, 1836) , Entomobryoides purpurascens ( Packard, 1873) , Willowsia buskii ( Lubbock, 1870) , Pseudosinella alba ( Packard, 1873) and Cyphoderus albinus Nicolet, 1842 ( Szeptycki 1979) .
Three specimens that are as large as the two first-instar specimens described below carry secondary scales and already have the reduced number of elements present in adults. Thus, Campylothorax is similar to Trogolaphysa in that the transformation and reorganisation of the chaetotaxy take place abruptly in the second-instar nymph, rather than the gradual change observed in Entomobryidae ( Soto-Adames 2015) .
Head
First-instar C. sabanus carries the eight rows of chaetae typically found in Entomobryoidea ( Figure 1 View Figure 1 A), but the number of elements is reduced when compared to Homidia , Seira , Pseudosinella and Trogolaphysa . As in Trogolaphysa , C. sabanus lacks chaetae M0 and S0, but in addition row M lacks M1, row Pa lacks Pa2 and row Pp is reduced to chaetae Pa1 and Pa3. The organisation of row M, with the three remaining elements pushed towards the eye patch, is a unique feature of Campylothorax .
The dorsal chaetotaxy of the head is further reduced in adults ( Figure 1 View Figure 1 B), where chaeta Ps3 is transformed into a scale and the only remaining elements in the posterior rows are Mc Pa5 and bothriotrix Pa6.
Mesothorax
The chaetotaxy of C. sabanus includes the same number of chaetae as in all other species of Entomobryoidea reported to date ( Figure 1 View Figure 1 C): three rows with 7, 6, 6 chaetae, respectively, and two lateral sensilla, one S-chaeta and one S-microchaeta. Scales are absent. The insertion of chaeta p2 is atypical in that it is displaced anteriorly towards m4, but otherwise most chaetae show the same organisation as in the first instar of other species.
The chaetotaxy in adults is markedly different from that in the first instar ( Figure 1 View Figure 1 D). In adults, elements in row a form the collar and all primary chaetae in row m are transformed into scales. In the posterior row, chaetae p1 – p4 are incorporated in the p3 Mc complex, whereas primary chaetae p5 and p6, and secondary chaeta p6e, remain as singletons.
Metathorax
The primary chaetotaxy includes 16 chaetae, one scale and one lateral S-chaeta ( Figure 1 View Figure 1 C). Adult C. sabanus retains four inner Mc in the p2 – 3 complex ( Figure 1 View Figure 1 D). The unique columnar arrangement of elements in the p2 – 3 complex allows for two possible interpretations of their identity. Assuming primary elements are conserved between Trogolaphysa and Campylothtorax, the anterior Mc may correspond to a2 and a4 ( Soto-Adames 2015). The slight internal displacement of the second anterior element suggests it is a displaced a2. This interpretation is supported by the similar arrangement of elements in Trogolaphysa and C. mitrai Bellini & Maneses, 2012 . Alternatively, comparison with C. sabanus first-instar chaetotaxy suggests that in adults, element a2 is transformed into a scale and the anterior Mc correspond to a4 and a5.
First abdominal segment
The primary chaetotaxy includes 11 – 12 chaetae, one scale and one S-microchaeta ( Figure 1 View Figure 1 E). This is the only species known to carry seven primary posterior (including rows m and p) chaetae ( Szeptycki 1979). The chaeta inserted external to the S-microchaeta (arrow in Figure 1 View Figure 1 E) is present only in one individual and it is likely spurious. The four elements retained in adults ( Figure 1 View Figure 1 F) are consistent with the nomenclature applied by Soto-Adames et al. (2014).
Second abdominal segment
The primary chaetotaxy in C. sabanus is complete, when compared to other entomobryoids, and includes two bothriotricha, 14 chaetae, one scale and one S-chaeta ( Figure 2 View Figure 2 A). Adults retain the two bothriotricha, two Mc, S-chaeta and four external microchaetae ( Figure 2 View Figure 2 B). The reduction in the number of inner chaetae in adults could be accounted for by the putative transformation of some elements into fan-shaped chaetae and their simultaneous displacement towards the bothriotricha. In this scenario, fan-shaped chaetae associated with bothriotrix m2 could account for elements a1, a2 and a3, whereas the chaetae associated with bothriotrix a5 would be p4, m4, a6 and m3e. I suggest that the additional chaeta associated with bothriotrix a5 corresponds to secondary chaeta m3e, which is present in almost all adult entomobryoids studied so far. Alternatively, all fan-shaped elements associated with the bothriotricha may be secondary (hence the question mark in Figure 2 View Figure 2 B) and the conserved number of chaetae is unrelated to primary chaetotaxy.
Third abdominal segment
The primary chaetotaxy of Campylothorax includes 14 chaetae, one scale, one S-chaeta, one S-microchaeta and three bothriotricha ( Figure 2 View Figure 2 C). The chaetae inserted between m4 and p5 (arrow) are present only on one side of one specimen. One unique characteristic of C. sabanus is the insertion of a3 internal to a2. The inverted insertion pattern between a2 and a3 has not been reported in the primary chaetotaxy of other species, but it is common in adults of the genus Lepidocyrtus ( Christiansen and Bellinger 1992; Soto-Adames 2000).
Adults have most chaetae transformed into fan-shaped microchaetae or Mc ( Figure 2 View Figure 2 D). As with Abd. 2, the fan-shaped elements associated with bothriotrix m2 may represent a2 and a3. The number of supplementary elements associated with bothriotricha a5 and m5 makes attempts at determining homology arbitrary and futile. The three mesochaetae or small Mc lateral to the bothriotrichal complex are interpreted as being primary, in series 7, whereas the inner microchaetae are considered secondary (a7i and p7i).
Fourth abdominal segment
The primary chaetotaxy of columns A, B, C, T, D, E and F comprises 4, 5, 4, 6, 3, 4, 3 elements distributed as in Figure 3 View Figure 3 A. In total, first-instar C. sabanus carries two scales, six Mc, two bothriotricha (inserted anterior to the pseudopore), 19 finely ciliate chaetae, S-chaeta posterior sensillum (ps) associated with T7 and 14 other S-chaetae inserted in the medial field bound by the bothriotrichal complexes. The medial S-chaetae are asymmetrically distributed, were not studied in detail and are not included in Figure 3 View Figure 3 A. The spatial distribution of most elements approximates the model described for T. jataca ( Soto-Adames 2015) . The homology of elements B1 and B6 is less clear. The anteriormost element in column B may be either B1 or B2; here this scale is identified as B1, assuming conservation in element homology in comparison with Trogolaphysa . The element between columns B and C is identified as B6 because its latero-external displacement and spatial relationship with elements B5, C3 and C4 is almost identical to that reported by Szeptycki (1979) for first-instar O. fl avescens. Campylothorax sabanus and O. fl avescens are the only two species known to show this peculiar displacement of element B6.
The anterior and posterior displacement of inner elements in adults make the determination of their homology a matter of speculation. One element is inserted near the anterior margin, but most other elements are displaced and grouped together near the posterior margin of the segment ( Figure 3 View Figure 3 B, C); even primary bothriotricha T2 and T4 are inserted posterior to the psedudopore. The homologies proposed below ( Figure 3 View Figure 3 B, C) are based on the principle of conservation of primary elements, as stated in the Material and methods section, and may be summarised by indicating that if the number and relative spatial distribution of primary elements is retained in the adult, then the elements present in adults are the same as in first-instar nymphs ( Szeptycki 1979).
Thus, the anterior paired Mc are identified as A1, whereas elements A3, A5, A6, B3, B4 and B5 are pushed towards the posterior margin and retained as or transformed into Mc. Element B6 moves laterally into column C ( Figure 3 View Figure 3 C) and develops into the medioposterior bothriotrix. Primary chaeta T6 is further displaced latero-externally. Except for element E1, all other lateral primary elements in columns E and F are transformed into Mc.
As indicated above, the great displacement of elements complicates homology determination, and alternative hypotheses are possible. Figure 3 View Figure 3 D shows one such alternative hypothesis of homology for all posterior and lateral Mc on Abd. 4. In this interpretation, the anterior inner element (not shown on Figure 3 View Figure 3 D) corresponds to A3 and the inner posterior cluster develops following the duplication of elements A5 and B5. In Figure 3 View Figure 3 D, the lateral chaetotaxy includes duplications of elements E3 and F1, and the latero-posterior bothriotrix corresponds to F3.
In adults two S-chaetae are easily identified, anterior sensillum (as) inserted close to C2 and ps inserted close to T7 ( Figure 3 View Figure 3 B – D). The medial region of the segment retains 12 – 14 S-chaetae present in the first instar, but their distribution was not studied and they are not included in Figure 3 View Figure 3 C – D.
The most prominent secondary elements added in adults are the medio-anterior Mc and the latero-posterior bothriotrix ( Figure 3 View Figure 3 B – D). The latero-posterior bothriotrix may be provisionally identified as F3p (or F3), but its true homology is unclear. A medial column of finely cilliate microchaetae is present in all adults examined (shown in fig. 62 in Soto-Adames et al. 2014), but these chaetae are short and difficult to see in most individuals. Other secondary elements appear to be variable. The two posterior medial Mc in Figure 3 View Figure 3 B (arrows) are modified elements of the medial column of microchaetae. The specimen with three medial Mc was collected in sympatry with two other individuals that carry only the anteriormost Mc. Other individuals examined carry supernumerary (usually one) Mc associated with the inner posterior group.
The most important differences between the present homology interpretation and that of Soto-Adames et al. (2014) concern the identity of microchaeta T6, the medioposterior bothriotrix and the inner posterior Mc, as shown in Table 1 View Table 1 .
Fifth abdominal segment
First-instar C. sabanus carries eight microchaetae, three Mc and three S-chaetae ( Figure 3 View Figure 3 D); there is no evidence of pseudopores in any of the juveniles or adults examined. The number and arrangement of most chaetae are similar to those described for other entomobryoids. The number and distribution of S-chaetae are identical to those of Sinella umesaoi Yosii, 1940 ( Zhang and Deharveng 2015) in that the lateral sensillum is inserted in the posterior row and internal to p5. In H. jordanai , the lateral sensillum is inserted in the middle row, near m5, whereas in all other species with three S-chaetae, the lateral sensillum is inserted external to m5/p5.
The adult chaetotaxy adds five secondary elements ( Figure 3 View Figure 3 E). The homology of secondary elements internal to m5 is consistent with those in other entomobryoids, but the affinities of elements external to m5 are unclear. The number and identity of secondary elements in Campylothorax are similar to those in Trogolaphysa . Differences between the two genera are the presence of p3pi and p6ae in Campylothorax and their absence in Trogolaphysa , whereas element m3a is present in Trogolaphysa but absent in Campylothorax .
In most individuals examined for the taxonomic analysis below, Abd. 5 is imbricated under Abd. 4 and only the Mc are clearly visible. In all the material from Hispaniola where the complete chaetotaxy of Abd. 5 is visible, it is identical to that of adult C. sabanus .
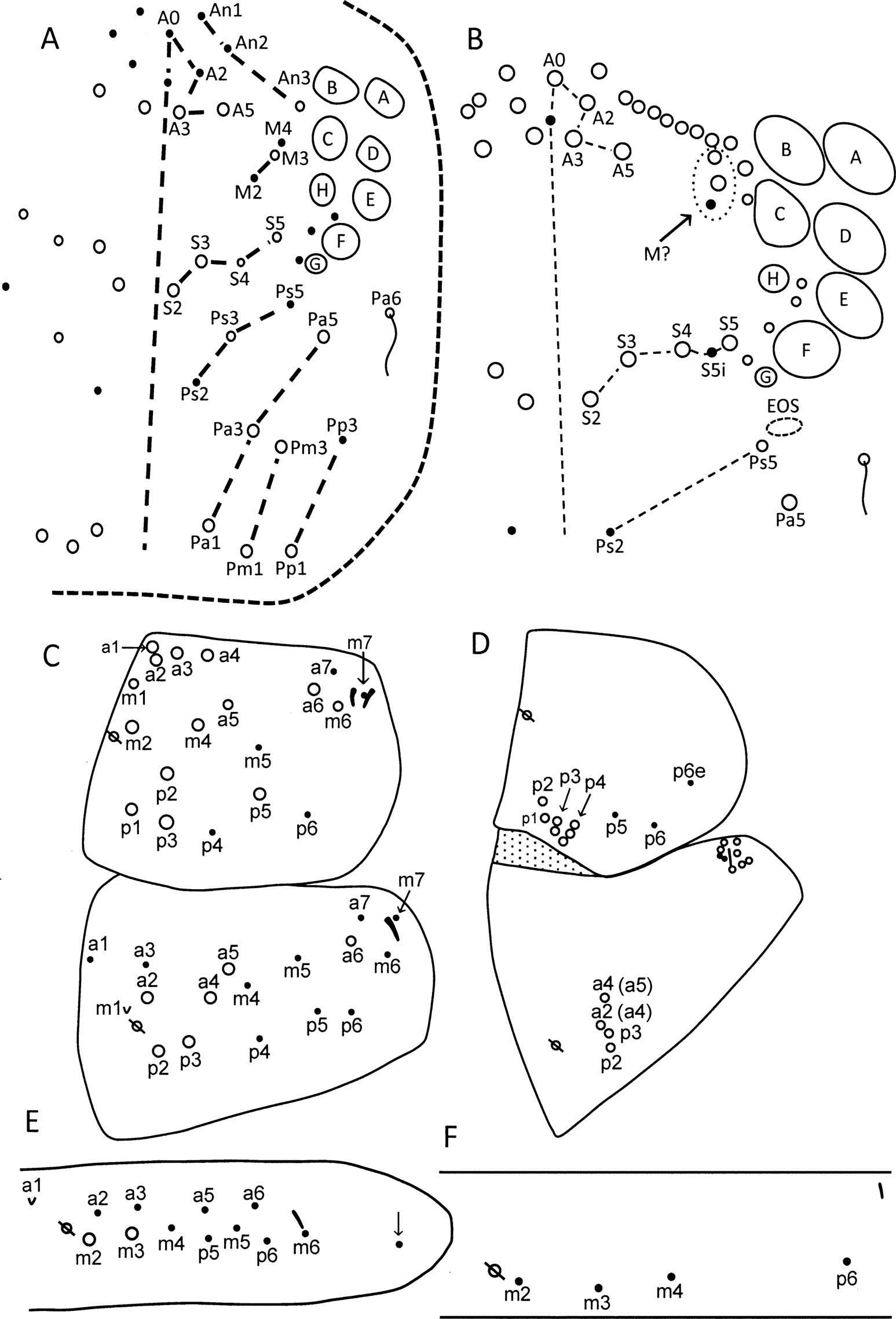
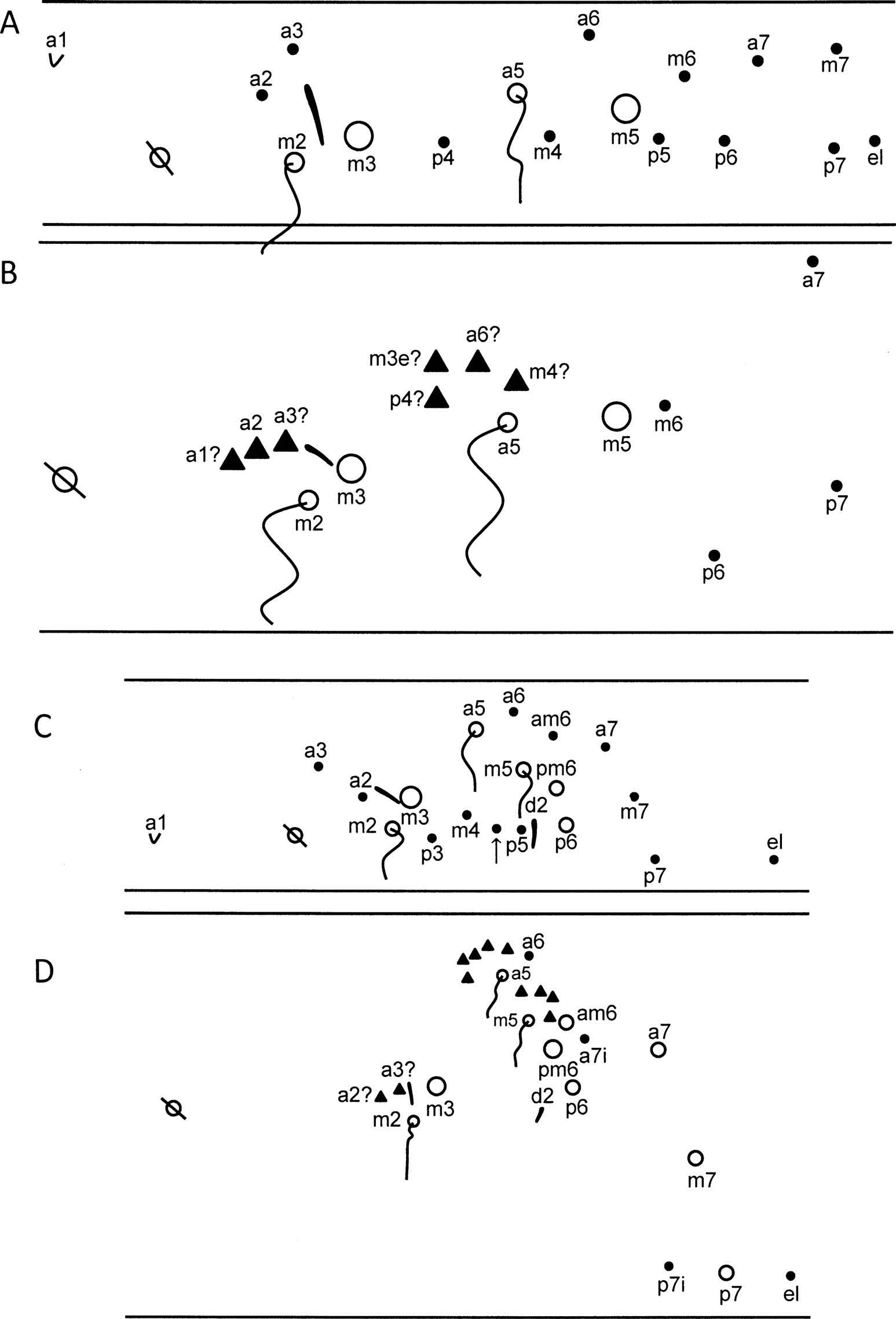
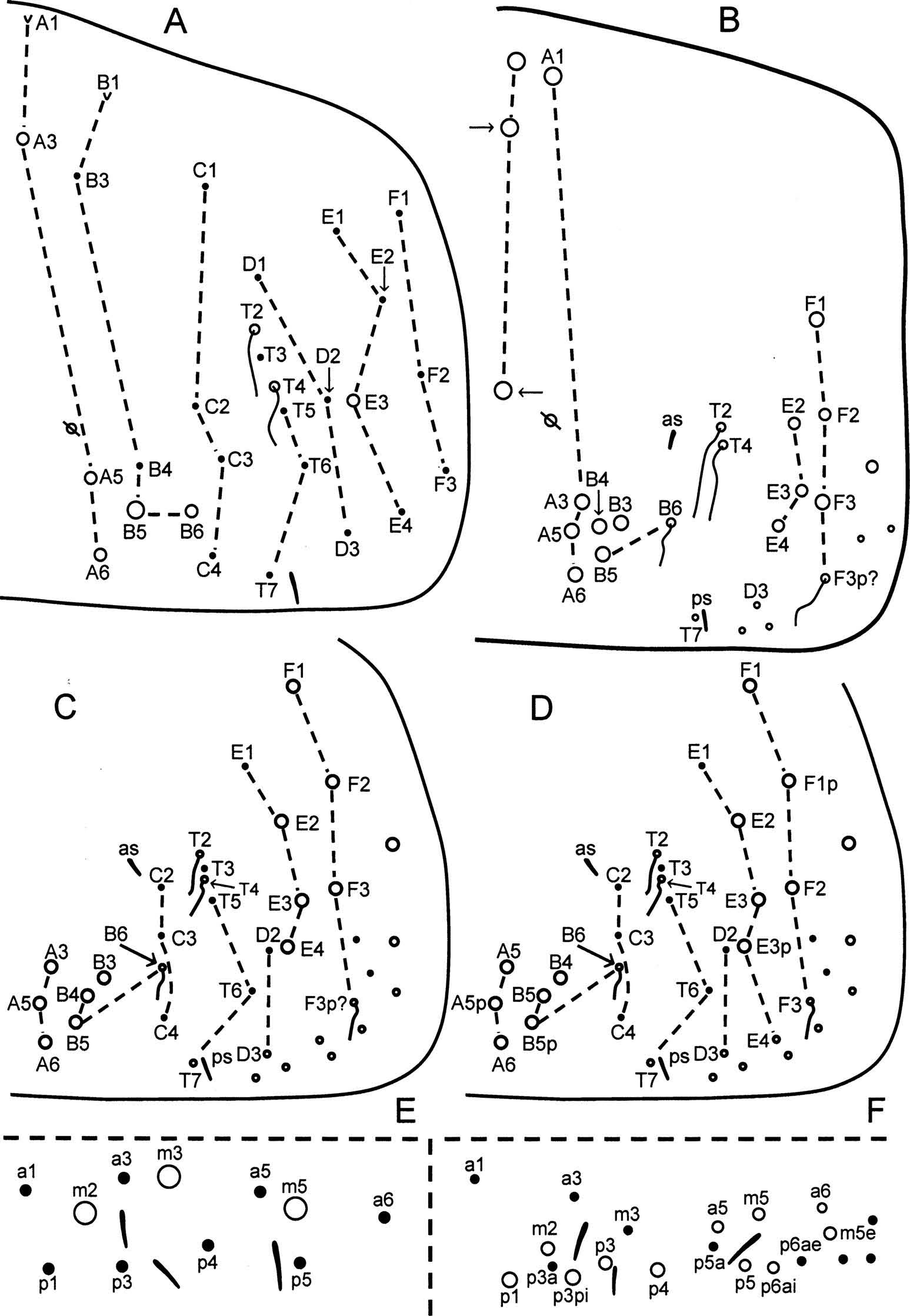
Table 1. Correspondence of selected elements of the fourth abdominal segment presented here, and those of Soto-Adames et al. (2014). Element designations in parentheses refer to alternative homology proposed in Figure 3 D.
| Present study | Soto-Adames et al. (2014) |
|---|---|
| A3 (A5) | A4 |
| B3 (B4) | B4 |
| B4 (B5) | B5 |
| B5 (B5p) | B6 |
| B6 | T6 |
| T6 | Unlabeled |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |