
Monatractides alticolus, Pešić, Vladimir & Smit, Harry, 2011
|
publication ID |
https://doi.org/10.5281/zenodo.207512 |
|
DOI |
https://doi.org/10.5281/zenodo.6188346 |
|
persistent identifier |
https://treatment.plazi.org/id/03F1878D-DC47-D839-F9F4-C0E0CDEEFCBC |
|
treatment provided by |
Plazi |
|
scientific name |
Monatractides alticolus |
| status |
sp. nov. |
Monatractides alticolus sp. nov.
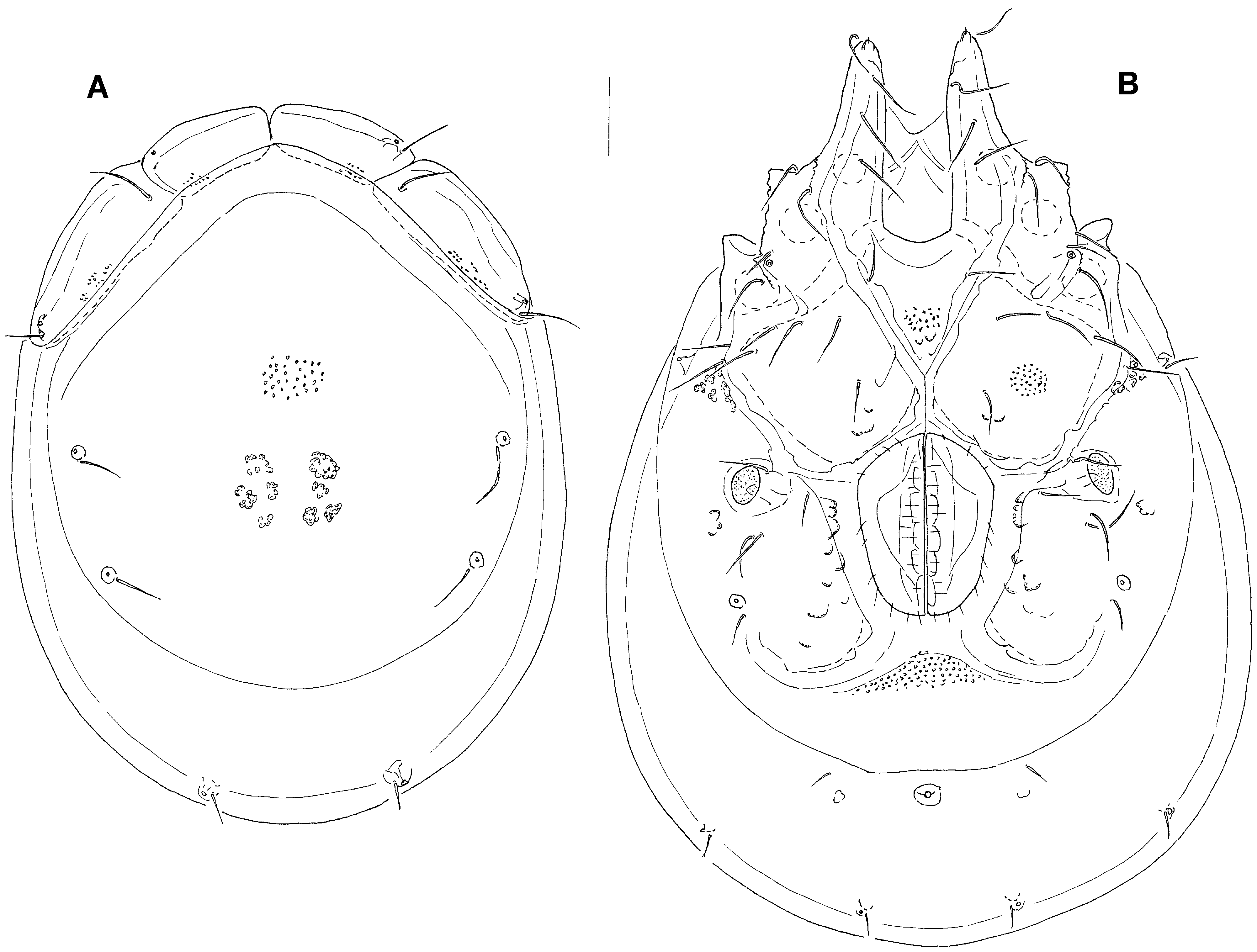
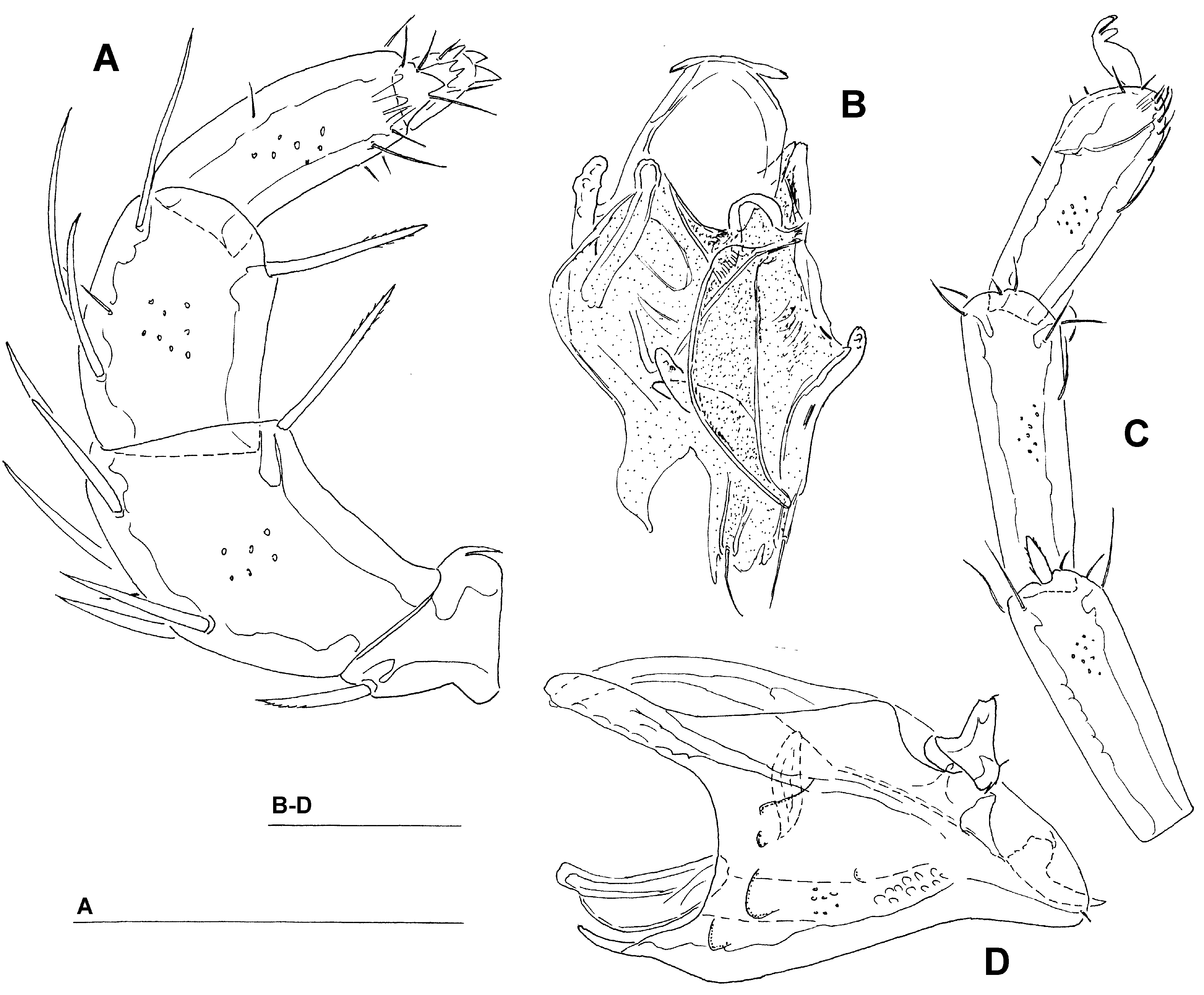
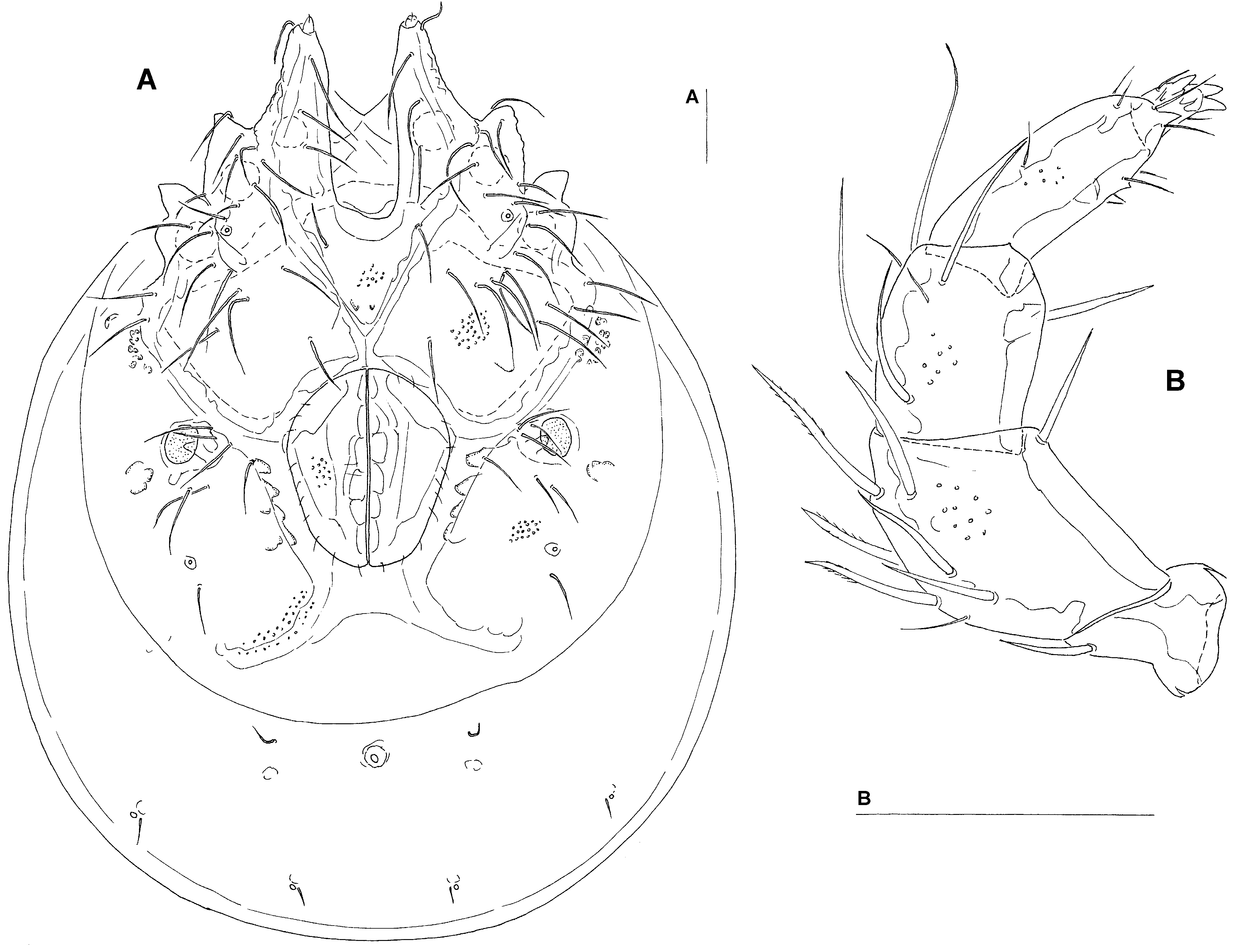
( Figs. 14 – 16 View FIGURE 14 A – B View FIGURE 15 A – D View FIGURE 16 A – B )
Type series. Holotype, male, dissected and slide mounted; Indonesia, New Guinea, Papua Province: Bion River, upstream, Pass Valley, 15.iii.2010, 3˚ 51.84 S 139˚ 05.733 E, 1962 m asl. Paratypes: 1/1 (0/1 mounted), same data as holotype; 2/2 (1/1 mounted), same data as holotype, 15.iii.2010, 3˚ 51.513 S 139˚ 05.570 E, 2007 m a.s.l..; 3/1, same data as holotype, 02.iii.2010, 3˚ 51.267 S 139˚ 05.789 E, 1962 m a.s.l.
Further records: Indonesia, New Guinea, Papua Province: Wao & Pao streams near confluence Walmak, Nipsan district, 18.iii.2010, 4˚ 07.097 S 139˚ 38.520 E, 1739 m a.s.l. 1/0 (mounted); Yango stream, Walmak, Nipsan district, 19.iii.2010, 4˚ 06.830 S 139˚ 38.587 E, 1757 m a.s.l. 2/2 (1/0 mounted).
Diagnosis. Idiosoma L> 1100 in males,> 1200 in females; area of primary sclerotization of the dorsal plate with four dorsoglandularia; capitular bay narrow (L/W ratio ca. 3.0); distal margins of P-2 and -3 without denticles, P-4 with small denticle near the insertion of the ventral hairs, P-4 ventral hairs inserted more distally, P-4 L/H 2.5 – 2.6; postgenital area large; excretory pore away from the line of primary sclerotization, Vgl-2 posterior and well separated from the excretory pore.
Description. Male ( holotype, in parentheses paratype from Bion River, in square brackets specimen from Wao & Pao streams): Idiosoma: (ventral view: Fig. 14B View FIGURE 14 A – B ) L 1163 (1219) [1100], W 806 (900) [744]; dorsal shield ( Fig. 14A View FIGURE 14 A – B ) L 906 (996) [878], W 700 (781) [644], L/W ratio 1.29 (1.28) [1.36]; dorsal plate 860 (934) [825]; shoulder plate L 259 (278) [228], W 97 (109) [97], L/W ratio 2.7 (2.6) [2.4]; frontal plate L 166 (209) [178], W 71 (92) [81], L/W ratio 2.3 (2.3) [2.2]; capitular bay L 258 (256) [238], W 82 (81) [80], L/W ratio 3.15 (3.16) [3.0]; Cx-1 total L 416 (433) [392], Cx-1 medial L 157 (175) [153], Cx-2+3 medial 80 (70) [68]; ratio Cx-1 L/Cx-2+3 medial L 5.2 (6.2) [5.8]; Cx-1 medial L/Cx-2+3 medial L 2.0 (2.5) [2.25]; genital field L/W 238 (261) [219]/169 (184) [159], L/ W ratio 1.4 (1.4) [1.38], ejaculatory complex ( Fig. 15B View FIGURE 15 A – D ) L 266 (292) [225]; distance genital field–excretory pore 216 (259) [206], genital field–caudal idiosoma margin 409 (440) [409]; capitulum ( Fig. 15D View FIGURE 15 A – D ) ventral L 272 (275) [247]; chelicera total L 313 (314) [303]; palp ( Fig. 15A View FIGURE 15 A – D ) total L 292 (316) [294], dL: P-1, 39 (40) [39]; P-2, 87 (92) [85]; P-3, 66 (65) [60]; P-4, 74 (86) [79]; P-5, 26 (33) [31]; %L: P-1, 13.4 (12.7) [13.3]; P-2, 29.8 (29.1) [28.9]; P- 3, 22.6 (20.6) [20.4]; P-4, 25.3 (27.2) [26.9]; P-5, 8.9 (10.4) [10.5]; L P-2/P-4 ratio, 1.18 (1.07) [1.08]; L I-L-4-6 ( Fig. 15C View FIGURE 15 A – D ): 151 (169) [160], 162 (178) [157], 145 (159) [142].
Female (from Bion River, upstream, n = 2): Idiosoma (ventral view: Fig. 16A View FIGURE 16 A – B ) L 1263 – 1356, W 988 – 1000; dorsal shield L 1081 – 1109, W 850 – 856, L/W ratio 1.27 – 1.3; dorsal plate L 1013 – 1044; shoulder plate L 256 – 272, W 109 – 122, L/W ratio 2.1 – 2.5; frontal plate L 203 – 213, W 97 – 100, L/W ratio 2.03 – 2.26; shoulder/frontal plate L ratio 1.24 – 1.26; capitular bay L 266 – 279, W 86 – 91, L/W ratio 3.1; Cx-1 total L 431 – 466, Cx-1 medial L 164 – 184, Cx-2+3 medial 37 – 38; ratio Cx-1 L/Cx-2+3 medial L 11.3 – 12.3; Cx-1 medial L/Cx-2+3 medial L 4.3 – 5.0; genital field L/W 269 – 278/218 – 222, L/W ratio 1.2 – 1.28; distance genital field–excretory pore 248 – 306, genital field–caudal idiosoma margin 497 – 552; capitulum ventral L 291 – 294; chelicera L 328 – 335; palp ( Fig. 16B View FIGURE 16 A – B ) total L 330 – 331, dL and %L (in parentheses): P-1, 43 – 47 (13.0 – 14.2); P-2, 96 (29.0 – 29.1); P-3, 68 – 70 (20.6 – 21.1); P-4, 86 – 89 (26.0 – 27.0); P-5, 32 – 34 (9.7 – 10.3); P-2/P-4 ratio, 1.08 – 1.12; L I-L-4-6: 174 – 175, 182 – 188, 162 – 169.
Discussion. Monatractides alticolus sp. nov. and M. humilis sp. nov. (see below) belong to M. macroporus ( Viets, 1935) species-complex (see Pešiċ & Smit 2009). Due to the less narrow capitular bay (L/W 3.0), larger idiosoma and gnathosoma dimensions and the presence of four dorsoglandularia in the area of primary sclerotization of the dorsal plate, the specimens from New Guinea are close to M. major ( Viets, 1935) , a species described originally from Java as a form of M. macrognathus ( Viets 1935) . Lundblad (1971) considered M. macrognathus and M.
macrognathus major , synonymous with M. macroporus , but Wiles (1991) raised M. macrognathus major in ranking to a full species.
Both species of the M. macroporus ( Viets, 1935) species-complex from New Guinea, M. alticolus sp. nov. and M. humilis sp. nov. (see below) differ from M. major (and the other members of the macroporus -complex from Java, see Viets 1935) in Vgl-2 shifted far away from the excretory pore.
Monatractides australicus ( Cook, 1986) , the Australian member of the M. macroporus species-complex, has a similar less narrow capitular bay, as well as the presence of the four dorsoglandularia in the area of primary sclerotization of the dorsal plate (see: Cook 1986). Both species from New Guinea can be distinguished in having less broad frontal platelets and a proportionally much longer genital field. Monatractides alticolus sp. nov. differs from M. humilis sp. nov. in its major idiosoma and gnathosoma dimensions (e.g., idiosoma L> 1100 in males,> 1200 in females vs. idiosoma L < 900 in males, < 1000 in females in M. humilis sp. nov.). Further differences are found in a longer ejaculatory complex (compare Figures 15B View FIGURE 15 A – D and 17D View FIGURE 17 A – G ) and P-4 is somewhat more slender (L/H 2.5 – 2.7 vs. 2.2 – 2.3 in M. humilis ) with a slightly less pronounced denticle near insertion of the ventral hairs which are inserted more distally in M. alticolus .
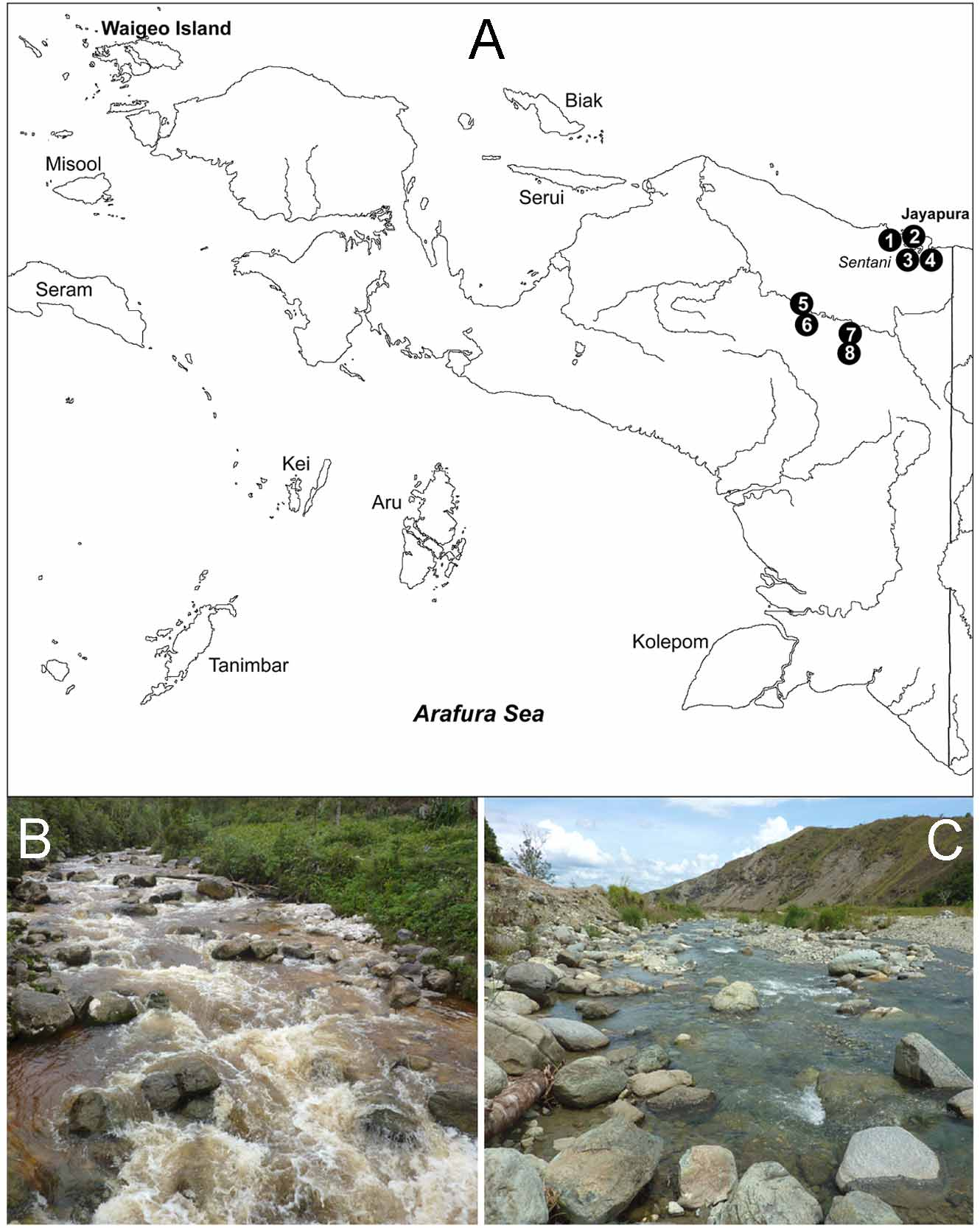
Apart from these diagnostic features, there may be ecological differences between these two species: Monatractides humilis sp. nov. was found only in lowland streams ( Fig. 1C View FIGURE 1 A – C. A ), while M. alticolus sp. nov. was restricted to higher elevations ( Fig. 1B View FIGURE 1 A – C. A ).
Etymology. Named for its occurrence at higher elevations.
Distribution. New Guinea.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Monatractides |
Monatractides alticolus
| Pešić, Vladimir & Smit, Harry 2011 |
Monatractides australicus (
| Cook 1986 |