
Sefapanosaurus zastronensis, Otero & Krupandan & Pol & Chinsamy & Choiniere, 2015
|
publication ID |
https://doi.org/10.1111/zoj.12247 |
|
publication LSID |
lsid:zoobank.org:pub:24B94C08-B94A-4B66-84F3-96C2F27E1FC6 |
|
persistent identifier |
https://treatment.plazi.org/id/03F187AE-E92A-2D3A-DF26-F916144EFEDD |
|
treatment provided by |
Felipe |
|
scientific name |
Sefapanosaurus zastronensis |
| status |
|
SEFAPANOSAURUS ZASTRONENSIS GEN. ET SP. NOV.
Holotype
BP/1/386, incomplete articulated left pes including astragalus, calcaneum, a putative distal tarsal IV, proximal portions of metatarsals III and IV, and almost complete metatarsal V.
Referred material
BP/1/7409–7455. Partial skeletons of at least four individuals comprising: BP/1/7409, a cervical vertebra; BP/1/7410, 7411, 7412, cervical centra; BP/1/ 7413, a partial cervical centrum?; BP/1/7414, middle dorsal vertebra; BP/1/7415–7417, three partially preserved dorsal vertebrae; BP/1/7418, a dorsal transverse process; BP/1/7419, a dorsal centrum; BP/1/ 7421, a posterior dorsal centrum; BP/1/7423, a posterior partial dorsal vertebra; BP/1/7431, an isolated dorsal neural arch; BP/1/7420, a dorsosacral centrum; BP/1/7422, a putative caudosacral vertebra; BP/1/ 7424–7429, six caudal vertebrae; BP/1/7430, an haemal arch; BP/1/7432, left coracoid; BP/1/7433, right scapula; BP/1/7434, proximal half of right humerus; BP/1/ 7435, right radius; BP/1/7436, right (?) radius; BP/1/ 7437, left ulna; BP/1/7438, incomplete articulated left manus consisting of distal carpals I and II, metacarpals I and II and phalanx I.1, and isolated manual digit V; BP/1/7439, proximal end of pubis (?); BP/1/7440–7443, proximal end of four femora; BP/1/ 7444, distal portion of femur; BP/1/7445, proximal end of right tibia; BP/1/7446, distal end of right fibula (?); BP/1/7447, left fibula; BP/1/7448, isolated proximal end of right metatarsal II; BP/1/7449, distal end of metatarsal III; and BP/1/7450, proximal end of ischium.
Information available at the Evolutionary Studies Institute (Johannesburg) collection database indicates that all the material described here (the holotype BP/1/386 as well as all the referred material) was ‘collected from the same locality’. These materials were catalogued as Euskelosaurus by James W. Kitching, and subsequently referred to Aardonyx by Adam Yates (a referral recently followed by McPhee et al., 2014). The referred material includes repeated elements, such as four partial femora of different sizes, indicating that this assemblage contains a minimum of four individuals. Despite the size differences, there are several autapomorphic features amongst the referred material that distinguish this assemblage from other sauropodomorphs (see Diagnosis). The available remains also lack autapomorphic features of other known basal sauropodomorphs and share characters typical of Anchisauria. Therefore, based on these features we deduce that there is no evidence to support the presence of more than one taxon amongst the material collected at this locality. The holotype is nonetheless restricted to the articulated remains of the tarsus and pes (BP/1/386), which were chosen based on the presence of autapomorphic traits that distinguish these elements from all other sauropodomorphs for which the tarsus and pes is known. The remaining material is catalogued under different collection numbers (BP/1/7409–7455) and referred here to the same taxon.
Type locality and horizon
The remains were collected by A. W. Keyser near the town of Zastron, in the Free State Province of South Africa, about 30 km west of the Lesotho border. There is no precise information about the exact stratigraphical level where the remains were found. However, around Zastron both lower and upper Elliot Formations crop out ( Bordy, Hancox & Rubidge, 2004); hence, we assign S. zastronensis to the Upper Triassic−Lower Jurassic Elliot Formation ( Fig. 1 View Figure 1 ).
Diagnosis
Sefapanosaurus zastronensis is a medium-sized basal sauropodomorph distinguished from other nonsauropod sauropodomorphs by the following autapomorphies present in the holotype: tall ascending process of the astragalus, being 35% of the mediolateral length of the astragalar body; T-shaped, triradiate cross-section of ascending process of the astragalus; and ascending process of the astragalus framed medially and caudally by well-developed, straight, thick ridges, which have subcircular cross-sections.
Additionally, the material referred to S. zastronensis (BP/1/7409–7455) differs from other nonsauropod sauropodomorphs by the following unique combination of characters (autapomorphies denoted with an asterisk): slit-shaped posterior dorsal neural canal; length of the base of the proximal neural spine greater than half the length of the neural arch; crosssectional shape of distal caudal centra squareshaped; long ridge extending from the craniodorsal margin of the coracoid to the coracoid foramen*; craniomedial process of the ulna twice as long as the craniolateral process and distally tapered*; presence of biceps tubercle and caudodistal tubercles of the radius; distal carpal I with proximally pointing tip on the palmar surface, giving a triangular shape in palmar view*; strongly concave medial margin of metacarpal I; ventral margin of phalanx I.1 well developed with a proximally pointing tip; metacarpal V longer than wide with flat proximal surface; first metacarpal short and wide (proximal width is more than 100% of the total length of the bone); craniomedial projection on the distal end of the fibula*.
Description
Etymology
From the Sesotho language sefapano, meaning ‘cross’, and from the Greek saurus, meaning ‘lizard’, in reference to the cross T-shaped ascending process of the astragalus. The specific name makes reference to Zastron, the type locality.
Axial skeleton
The major components of the entire axial column are all represented by at least one vertebra, although some elements have only preserved the centrum or the neural arch. There are 12 presacral vertebrae, an isolated neural arch, an isolated transverse process, a dorsosacral vertebra, a putative caudosacral vertebra, and six caudal vertebrae. The descriptions below were based mainly on the best-preserved elements, which present the neural arch.
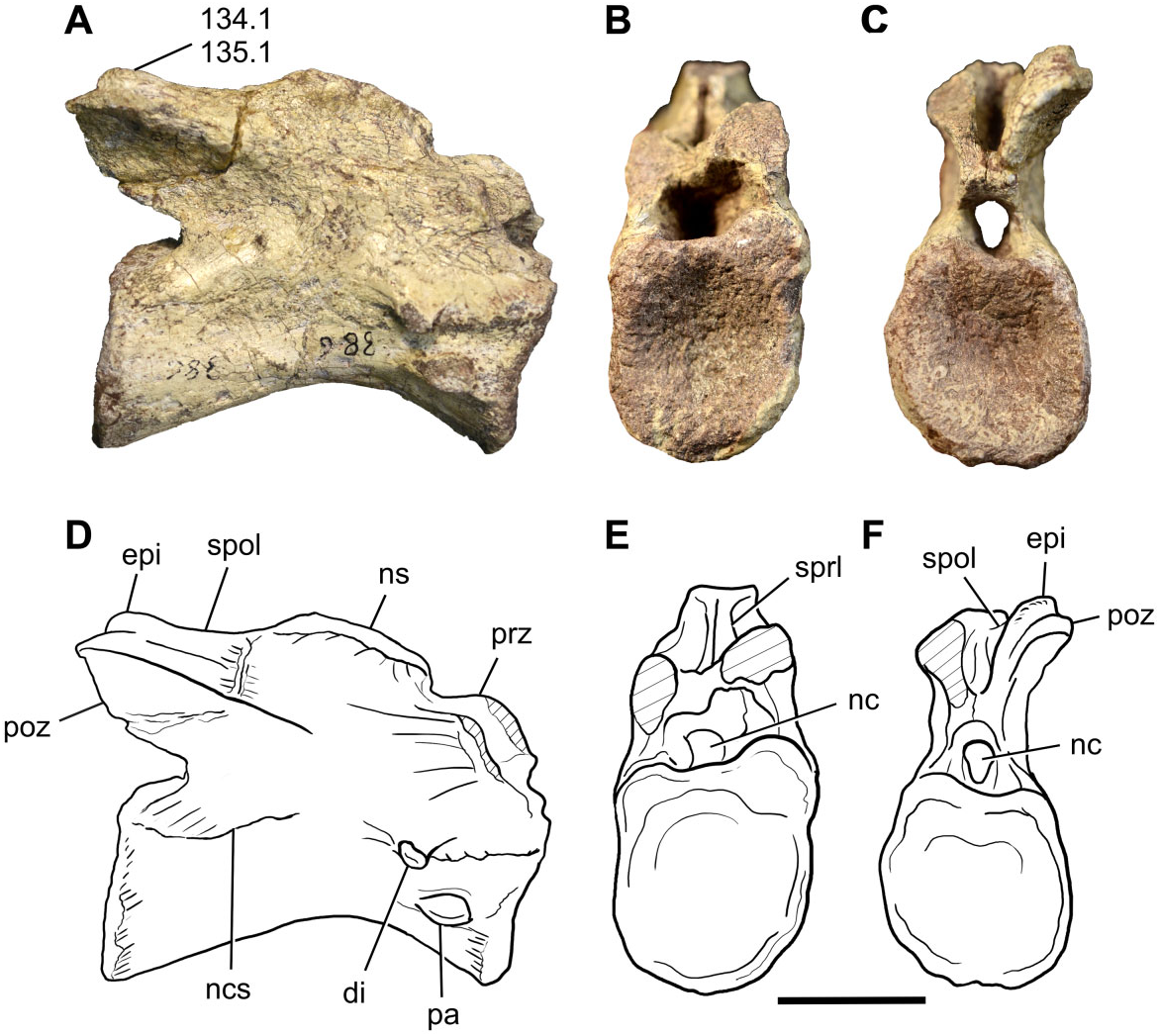
Cervical vertebrae: A nearly complete cervical vertebra (BP/1/7409) is preserved in Sefapanosaurus , plus four isolated cervical centra (BP/1/7410, 7411, 7412, 7413). The former is probably the fifth or sixth cervical vertebra ( Fig. 2 View Figure 2 ). The general morphology of the nearly complete vertebra is typical for a nonsauropodan sauropodomorph cervical element: it is elongate and low, the height of the neural arch is less than the height of the centrum, and there are no depressions on the lateral surface of the centrum ( Galton & Upchurch, 2004). The length of the centrum is 2.25 times the height of the cranial surface of the centrum, and the latter is 1.2 times the cranial face width (see Table S2 for measurements). A neurocentral suture is clearly visible, although obliterated in BP/1/7409. The presence of an isolated cervical centrum suggests that this element may belong to an immature individual ( Brochu, 1996; Irmis, 2007).
The articular facets of the centra are amphicoelous, the cranial surface being shallowly concave and the caudal face deeply excavated, as is the generalized condition for basal sauropodomorphs, and both the cranial and caudal articular facets are subequal in size. The ventral and lateral margins are concave. The ventral surfaces of the centra lack a keel, having only a low ridge on the cranial portion. Immediately caudal to this low ridge, several small foramina are present on BP/1/7409, but these foramina are not present in BP/1/7410. However, the ventral keel is a feature that is not always well developed along the cervical series amongst basal sauropodomorphs, being commonly present and more developed in the posterior cervical vertebrae (cervical eight to ten) and anterior dorsal vertebrae (dorsals one and two). This pattern is noticeable in the complete axial series of Plateosaurus longiceps ( MB. R. 4404) and Ruehleia bedheimensis ( MB. R. 4718). Other basal sauropodomorphs, however, have well-developed ventral keels along the entire cervical series (e.g. Leonerasaurus taquetrensis ).
The parapophyses are well developed and located at the midheight region of the lateral surface of the centrum. In lateral view the parapophyses are lacriform in shape, with a tapering posterior end and a rounded anterior end. This shape can only be seen on the right side of BP/1/7409, and the left side of BP/1/7410. Ventrally they have a sharply defined margin, visible in ventral view. The diapophyses are present, projecting lateroventrally in a pendant fashion, and are situat- ed on the cranioventral corner of the bases of the neural arches. This contrasts with the condition present in Aardonyx , in which the diapophyses are extremely reduced ( Yates et al., 2010). No diapophyseal laminae are present, as is common for midcervical vertebrae of sauropodomorphs more basal than Tazoudasaurus ( Galton & Upchurch, 2004) .
The prezygapophyses are not preserved in BP/1/ 7409, although both spinoprezygapophyseal laminae ( SPRL) are present. Only the right cervical postzygapophysis is complete. In posterior view, its mediolateral axis is orientated laterodorsally at an angle of approximately 30°, as commonly found in middle cervical vertebrae of most basal sauropodomorphs ( Galton & Upchurch, 2004). The caudal-most extent of the postzygapophysis exceeds the level of the caudal margin of the centrum. An epipophysis is present and well developed, extending along the entire length of the postzygapophysis and grading into the spinopostzygapophyseal lamina ( SPOL), as in Sarahsaurus , Lufengosaurus , and most sauropodiforms. The epipophysis does not reach the level of the caudal margin of the postzygapophysis, a condition also present in Pantydraco , Thecodontosaurus , Sarahsaurus , Leyesaurus , and Adeopapposaurus .
Only the base of the neural spine is preserved. The base of the spine is craniocaudally developed, as in the middle cervical vertebrae of most basal sauropodomorphs.
The isolated cervical centrum, BP/1/7010, is approximately the same length as that of BP/1/7409, but its rostral and caudal articular surfaces are about 10% mediolaterally wider and dorsoventrally taller. The parapophysis is of a similar shape and is in a topologically similar position to BP/1/7409. Comparisons with other basal sauropodomorphs (e.g. Adeopapposaurus , Plateosaurus engelthardti , Ruehleia ) suggest that it probably represents a middle cervical vertebra.
Dorsal vertebrae: Seven dorsal vertebrae are present in Sefapanosaurus , together with an isolated transverse process, and an isolated neural arch. Five elements (BP/ 1/7414, 7415, 7416, 7417, 7423) preserve partial or complete neural arches, two posterior elements lack neural arches (BP/1/7419, 7421), and there is also an isolated neural arch (BP/1/7431). The description is focused mainly upon the two better-preserved elements, which probably represent dorsal vertebrate five and ten. The isolated neural arch corresponds to the middle or midposterior region of the dorsal column series.
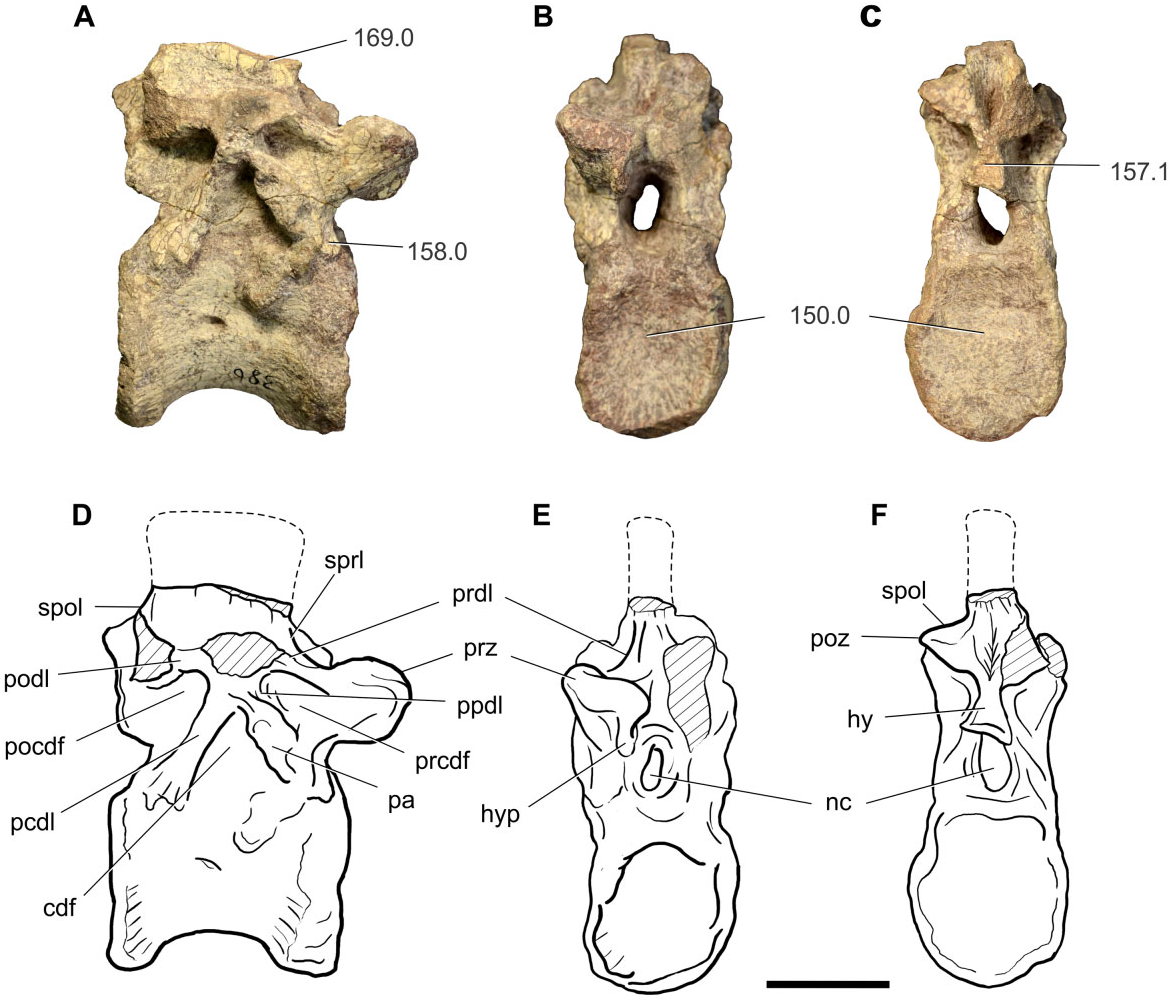
The anterior dorsal vertebra (BP/1/7415; probably the fifth dorsal) is almost completely preserved, in which the neural arch lamination and fossae are clearly visible ( Fig. 3 View Figure 3 ).
The centrum length is 1.2 times the cranial centrum height, and the latter is 1.3 times the width of the cranial facet (see Table S2 for measurements). Overall, the centrum is spool-shaped, with a constricted mesial region and expanded cranial and caudal ends. The cranial face is flat and caudal articular facets are shallowly concave, as is common for most basal sauropodomorphs. The facets are subcircular, being slightly taller than they are wide. The lateral surface bears a shallow fossa at approximately midheight immediately ventral to the neurocentral suture. The ventral surface is flat and lacks any sign of a ridge or keel, unlike the condition present in the last cervical and the anterior-most dorsals of most basal sauropodomorphs. The lateral surfaces of the centrum are also flat and no depressions are present. The centrum and neural arch are fused to each other, although a remnant line demarcating the neurocentral suture can be seen. Lateral surfaces of the centrum bear shallow foramina in their middle portions. The foramen on the right side opens caudally and is developed at the midheight of the centrum and slightly caudal to the anterior−posterior midline. The foramen on the left side is very weakly developed, and is located on the anterior−posterior midline but dorsally, close to the neurocentral suture.
The neural arch is low, not exceeding the centrum height, measured from the neurocentral suture up to the level of the zygapophyseal facets, a feature present in most nonsauropod sauropodomorphs. The neural spine is not completely preserved, but a craniocaudally elongated base can be seen. The distal ends of the parapophyses are eroded, but the preserved bases suggest that they are relatively large and ovoid in crosssection, with the long axis orientated caudodorsally. On the right side of the neural arch, a dorsoventrally elongated parapophysis defines most of the caudal boundary of the prezygapophyseal centrodiapophyseal fossa ( PRCDF) and the cranial boundary of the centrodiapophyseal fossa ( CDF) and is developed entirely on the neural arch. The PRCDF is also delimited by the prezygodiapophyseal lamina ( PRDL) and the paradiapophyseal lamina ( PPDL) and centrodiapophyseal lamina ( ACDL). The diapophyses are not completely preserved, but the proximal portion of the left diapophysis is present. Apart from the PRCDF and CDF, a postzygapophyseal centrodiapophyseal fossa ( POCDF) is also present. The CDF is delimited by the PPDL and the parapophysis cranially, and the posterior centrodiapophyseal lamina ( PCDL) caudally, whereas the POCDF is bounded by the postzygodiapophyseal lamina ( PODL) and the PCDL. Above the zygodiapophyseal table ( ZDT) only the spinodiapophyseal fossa ( SDF) can be recognized framed by the PRDL, SPRL, SPOL, and PODL. The preserved right prezygapophysis extends anterior to the level of the cranial margin of the centrum. The articular facet of the prezygapophysis has a slight dorsomedial inclination and bears a well-developed hypantrum on its ventromedial margin. The postzygapophyses bear SPOL dorsally, and a welldeveloped triangular hyposphene ventromedially, which does not exceed the dorsoventral height of the neural canal, as is common for all sauropodomorphs more primitive than Melanorosaurus ( Yates, 2007a) . The neural canal is dorsoventrally elongate, more than twice as tall as it is wide. It deeply excavates the dorsal surface of the centrum as a V-shaped furrow. Within this furrow, on the dorsal surface of the centrum, two shallow, lenticular foramina are present.
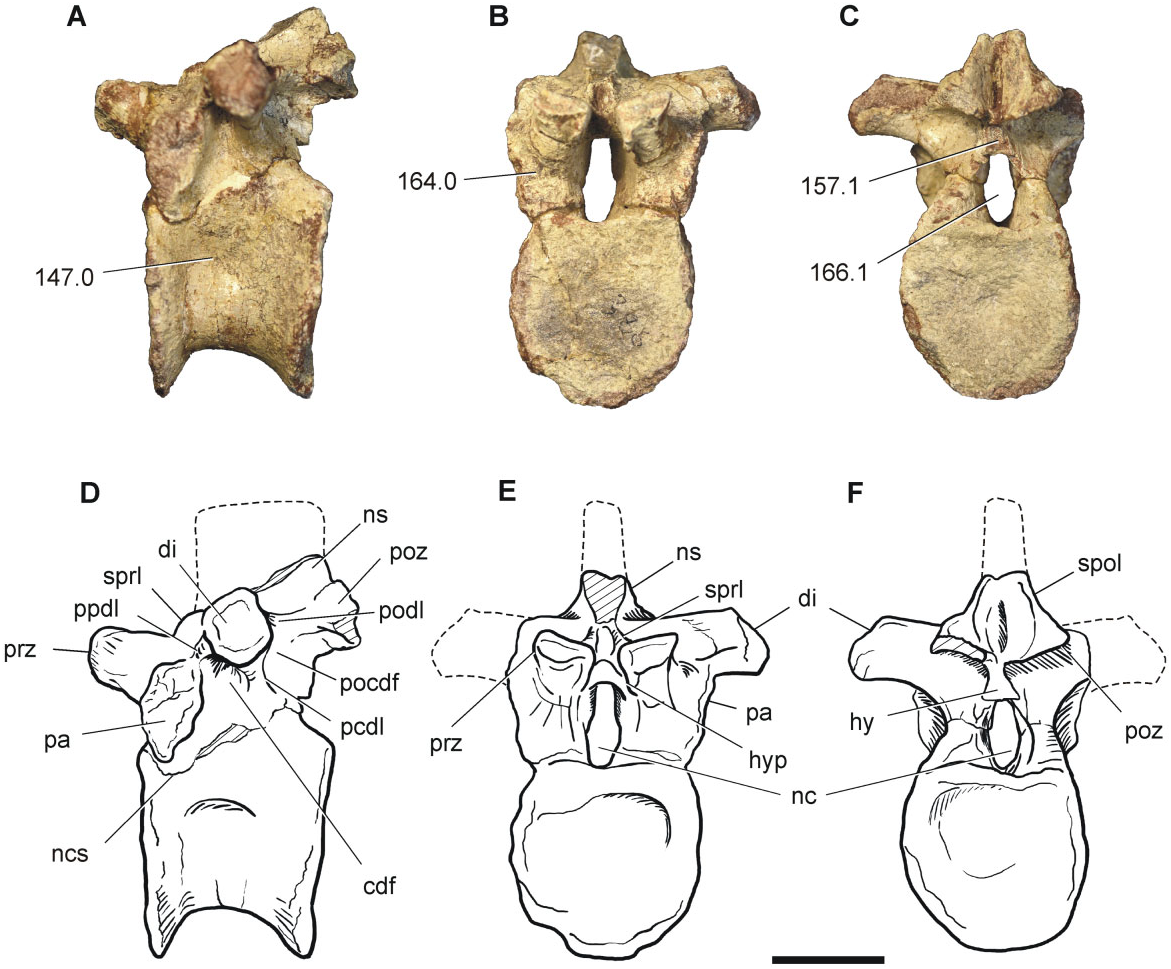
The middle dorsal vertebra (BP/1/7416, tenth or eleventh) is almost complete, lacking the right diapophysis and the neural spine ( Fig. 4 View Figure 4 ). The centrum is relatively short, with a length that is 0.85 times the cranial centrum height, and the latter being equal to the width of the cranial articular surface. The cranial and caudal articular facets are amphyplatyan to slightly amphycoelous, and the lateral surface of the centrum bears a shallow depression on its middle portion. The presence of dorsal vertebrae with vague or shallow depressions is a common feature amongst sauropodomorphs more basal than Isanosaurus , unlike the condition of sauropods, in which deep fossae or invasive pleurocoels are present ( Upchurch et al., 2007; Yates, 2007a). The contact between the centrum and the neural arch shows the trace of a neurocentral suture, although both structures are fused to each other.
As for the anterior dorsal vertebra, the neural arch is low, with a height that is less than that of the centrum. The parapophyses are well developed and entirely situated on the neural arch. The left diapophysis is preserved and is well developed laterally and almost horizontally orientated. A CDF is present below the latter lamina. The PCDL projects caudoventrally from the diapophysis and the PODL extends caudally, delimiting a POCDF. The PPDL extends cranioventrally. At this point of the axial column, no PRDL are present, a common feature of the middle dorsals of most basal sauropodomorphs ( Upchurch et al., 2007; Yates, 2007a; Pol et al., 2011). As a result, no PRCDF is present. The prezygapophyses exceed the cranial centrum margin and the angle between the two articular facets is approximately 165°. Hypantrum articulations are present and diverge from each other at an angle of approximately 35°. The postzygapophyses are situat- ed almost at the same level as the prezygapophyses, have subcircular articular facets, and slightly exceed the caudal margin of the centrum. Dorsally, the postzygapophyses are framed by SPOL. The hyposphenes are noticeable, although they are dorsoventrally shorter than the neural canal, as in nonsauropod sauropodomorphs.
Posterior dorsal vertebrae are represented by four centra (BP/1/7417, 7419, 7421, 7423), two of them (BP/ 1/7417, 7423) including the base of the neural arch. These elements present expanded cranial and caudal faces, which are circular in shape, and amphyplatyan to slightly amphicoelous. Shallow depressions are present on the lateral faces, close to the neurocentral suture, a feature common to most basal sauropodomorphs ( Galton & Upchurch, 2004). The element BP/1/7417 presents part of the ACDL and PCDL, which delimit the CDF.
Sacral vertebrae: Only one of the preserved vertebra can convincingly be attributed to a sacral element and corresponds to an isolated centrum that can be identified as a dorsosacral vertebra ( Fig. 5 View Figure 5 ). Besides this, a putative caudosacral vertebra is preserved.
The dorsosacral vertebra (BP/1/7420) has a centrum that is 0.95 times the cranial centrum height and the latter is 0.8 times the cranial centrum width. The centrum is constricted and bears a shallow depression at its middle region. The cranial portion of the centrum has the transverse process with the articulation for the sacral rib and extends from the neurocentral suture to the cranioventral margin. A similar pattern is present in the dorsosacral vertebra of Leonerasaurus ( Pol et al., 2011; fig. 8E) and in Melanorosaurus (NMQR 1551) .
The putative caudosacral vertebra (BP/1/7422), represented by an isolated centrum, is a robust element, with a centrum length that is 0.65 times the centrum height. The articular faces are amphiplatyan to slightly amphicoelous. Although the neural arch is not preserved, the base of the left transverse process does, showing a dorsoventral elongation, originating also from part of the centrum, as present in caudosacral vertebra of Plateosaurus engelthardti ( SMNS 91269).
Caudal vertebrae: Six caudal vertebrae are preserved, representing the anterior, middle, and posteri- or region of the tail ( Fig. 6 View Figure 6 ). The anterior-most caudal vertebra (BP/1/7424, probably the first) lacks the neural arch, only preserving part of the left transverse process. The centrum length is about 0.65 times the cranial centrum height, and the latter is almost the same length as the cranial centrum width. A short anterior caudal centrum is a feature present in Sauropodiformes. The articular facets are amphiplatyan to slightly amphicoelous. The lateral surface lacks depressions and the ventral surface has only traces of the chevron articular facets. There is no ventral longitudinal hollow. The transverse process preserved is not positioned on the neural arch, but in the dorsal region of the centrum, over its cranial half.
A second anterior caudal vertebra (BP/1/7425) is better preserved, with most of the neural arch, but preserving only the base of the neural spine. The cranial and caudal articular facets are slightly amphicoelous. The centrum length is 0.85 times the cranial centrum height, and the latter is 1.2 times its width. The ventral margin is strongly concave and the ventral surface bears a shallow longitudinal groove, which ends at the chevron articular facets. The centrum lateral surface lacks a depression. The transverse processes are dorsoventrally compressed, horizontally directed, and situated on the dorsal margin of the centrum, at the level of the neurocentral suture, as in most basal sauropodomorphs (except for Anchisaurus ). The neural arch lacks zygapophyses and the neural spine, preserving only the base of the latter, from which can be seen a portion of the SPRL. The length of the base of the neural spine is greater than half of the length of the neural arch, a feature unusual in most basal sauropodomorphs, except for basal-most forms (i.e. Chromogisaurus , Saturnalia , Thecodontosaurus, Efrassia ), ‘ Plateosaurus ’ gracilis , and the basal sauropod Tazoudasaurus .
A posterior caudal centrum has also been preserved (BP/1/7429). It is elongate, its centrum length being 1.5 times the cranial centrum height. The cranial and caudal articular facets are rounded and amphicoelous. The ventral and lateral surfaces are almost flat, giving the centrum a square-shaped crosssection, a character present in Adeopapposaurus and common for most transitional forms leading to Sauropoda (i.e. Anchisaurus , Mussaurus , Aardonyx , Melanorosaurus , and Camelotia ). The neural arch is missing, except for its base, which is situated cranially on the centrum. The transverse processes are broken.
Chevron: An isolated anterior chevron is preserved (BP/ 1/7430), lacking only the distal-most tip of the blade ( Fig. 7 View Figure 7 ). As is common for most basal sauropodomorphs, the chevron morphology closely resembles the closed ‘Y-shaped’ morphotype described by Otero et al. (2012) for sauropods.
Pectoral girdle and forelimbs
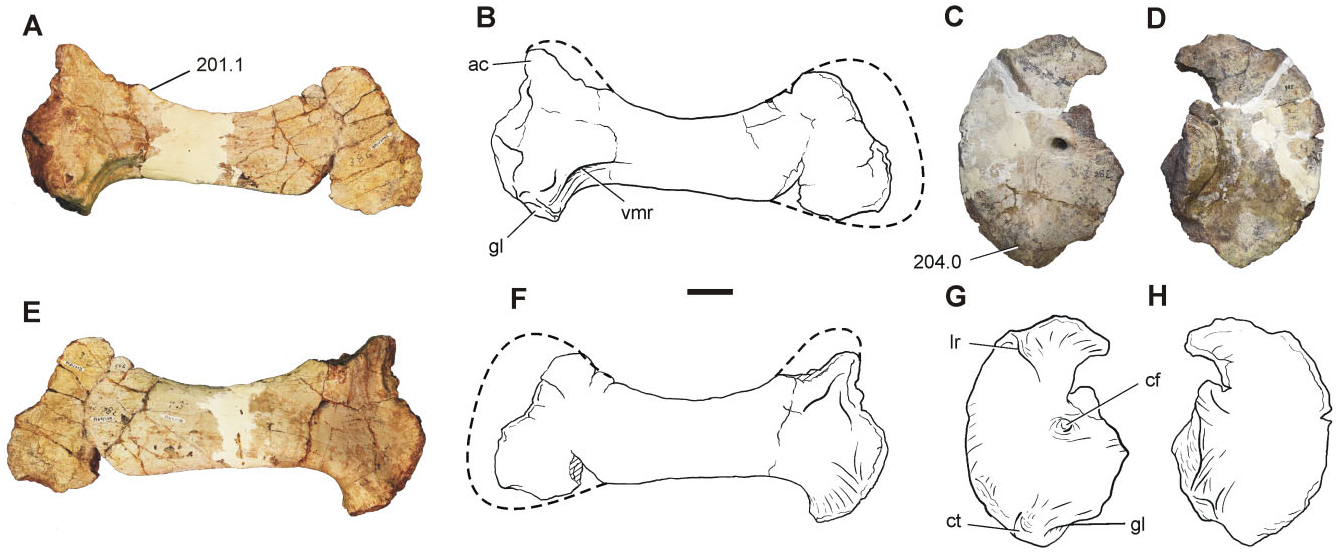
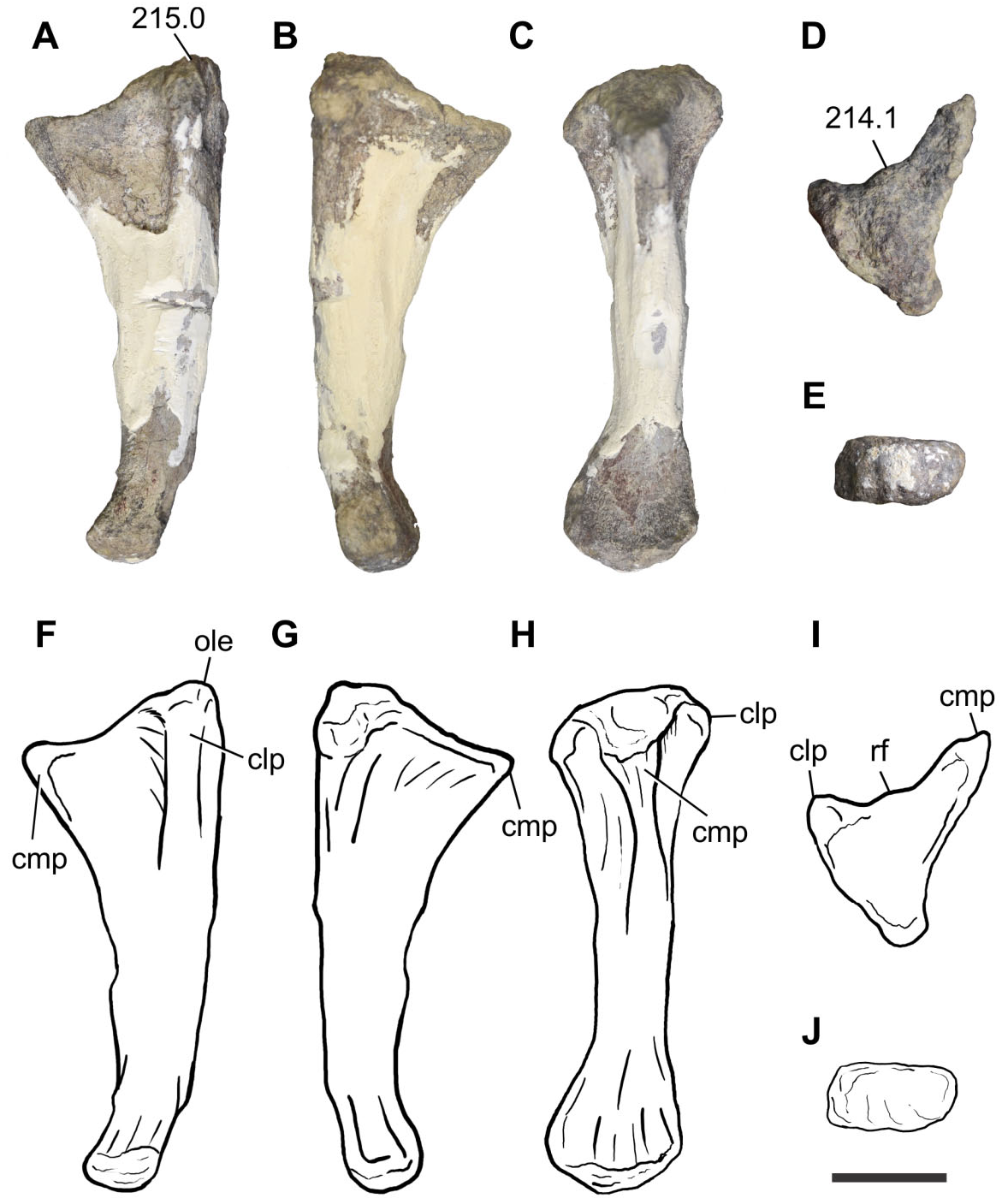
The preserved elements of the pectoral girdle include an almost complete right scapula (BP/1/7433) and left coracoid (BP/1/7432), whereas the forelimb bones are represented by a proximal portion of a right humerus (BP/1/7434), two radii (BP/1/7435 and 7436) and an ulna (BP/1/7437), and a partially articulated left manus (BP/1/7438), including distal carpals 1 and 2, metacarpals I, II, and V, and phalanges I-1, V-1, and V-2. Pectoral girdle: The right scapula (BP/1/7433) preserves most of its proximal portion, including part of the acromion process, as well as the majority of the blade ( Fig. 8A, B, E, F View Figure 8 ). Despite the fact that the acromion process is not complete, it can be determined that it rises from the scapular blade at an angle smaller than 65°, as in nonsauropod sauropodomorphs (with the exception of Saturnalia , Lufengosaurus, Jingshanosaurus , and Mussaurus ). The glenoid region is the thickest part of the scapula and the glenoid articular facet is subcircular. The ventral margin of the blade and the caudal margin of the glenoid project at an angle close to 90°, as in Coloradisaurus but differing from Plateosaurus , Adeopapposaurus , Mussaurus , and Euskelosaurus , which have an angle more than 90°. The medial surface of the scapula has a ventromedial ridge, a feature also present in Unaysaurus , Adeopapposaurus , Plateosauravus , Mussaurus , and Euskelosaurus , although only in Mussaurus is this feature extremely developed and reaches the distal third of the distal blade ( Otero & Pol, 2013). The minimum blade width is approximately 0.47 times the ventral expansion of the scapula. The general appearance of the scapula of Sefapanosaurus is similar to that of SAM-K386 assigned to Euskelosaurus ; however, the latter taxon differs from Sefapanosaurus by the presence of a more robust morphology, a stouter ventromedial ridge, and an angle between the ventral margin of the blade and the caudal margin of the glenoid close to 120°.
regarded here as an autapomorphic feature of Sefapanosaurus .
Humerus: Only the proximal portion of the humerus (BP/1/7434) is preserved ( Fig. 9 View Figure 9 ). This portion is mediolaterally expanded, as in all basal sauropodomorphs. Proximal abrasion of the bone precludes the identification of the internal tuberosity. The deltopectoral crest is subrectangular and rises from the proximal portion of the humerus and projects craniolaterally, as in most basal sauropodomorphs. The craniolateral margin is straight, as seen in most basal sauropodomorphs, with the exception of Coloradisaurus , Riojasaurus , Sarahsaurus , Plateosauravus , and Lessemsaurus , in which the crest is strongly sinuous in craniolateral view.
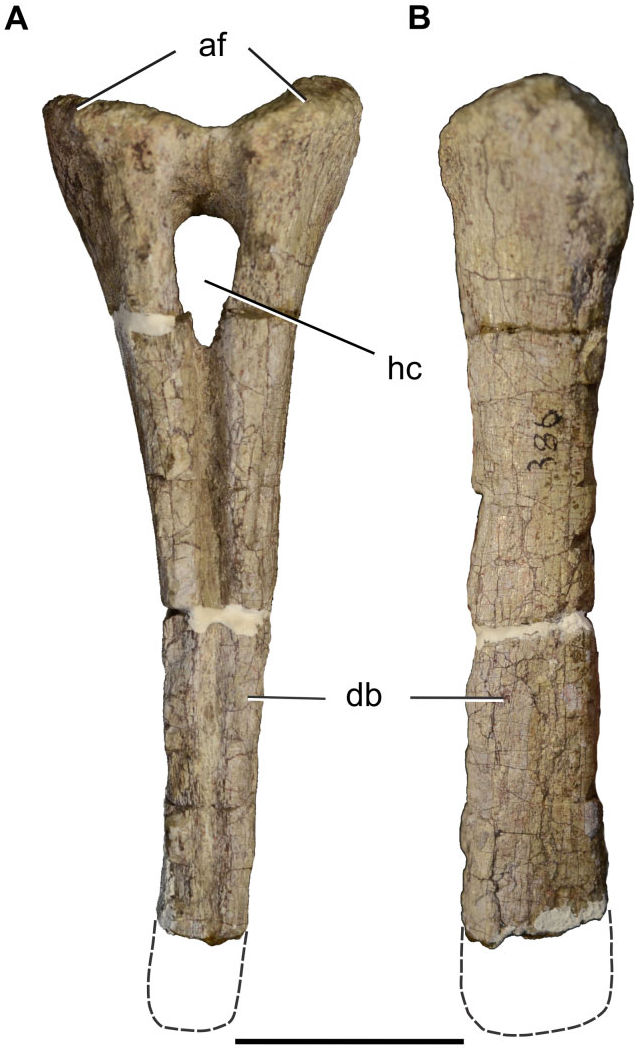
The left coracoid (BP/1/7432) is almost completely preserved, lacking only its caudodorsal portion ( Fig. 8C, D, G, H View Figure 8 ). The overall morphology is congruent with the generalized condition of most basal sauropodomorphs in being an ovoid bone with its major axis parallel to the suture with the scapula. The coracoid foramen is situated close to the caudal margin at the midlength of the coracoid. The ventral margin is the thickest and this thickness is given by the presence of a coracoid tubercle, a feature also present in Sarahsaurus , Adeopapposaurus , Coloradisaurus , and sauropods more derived than Tazoudasaurus . There is a noticeable long ridge extending from the craniodorsal margin of the coracoid to the coracoid foramen, although not reaching it. This feature has not been reported in other basal sauropodomorphs and so is Ulna: An almost complete left ulna (BP/1/7437) has been preserved ( Fig. 10 View Figure 10 ). The proximal end of the ulna is expanded both mediolaterally and craniocaudally, and has a triradiate profile in proximal view, given the development of the craniolateral and craniomedial processes. A triradiate proximal surface of the ulna is present in all sauropodomorphs more derived than Aardonyx . The craniolateral and craniomedial processes delimit a radial fossa, which is more developed in Sefapanosaurus than in Aardonyx , Mussaurus , and Melanorosaurus . The craniomedial process of the ulna is extremely developed, a condition also report- ed in Antetonitrus and Vulcanodon . Moreover, the craniomedial process of Sefapanosaurus tapers distally, unlike the condition in Mussaurus , Aardonyx , Melanorosaurus , and Antetonitrus . In the latter taxa, the craniomedial process of the ulna terminates in a more rounded fashion. The maximum proximal length (measured along the craniomedial process) is 0.42 times the total length of the ulna, whereas the minimum proximal length (measured along the craniolateral process) is 0.32 times the total ulnar length. The opposite margin (medial) of the radial fossa is also concave, a condition also present in other basal sauropodomorphs such as Melanorosaurus readi (SAM-PK-K3449) and the basal sauropods Lessemsaurus sauropoides ( Pol & Powell, 2007b) and Antetonitrus ingenipes ( BPI /1/ 4952), but absent in other sauropodomorphs (e.g. Vulcanodon , Yunnanosaurus , Tehuelchesaurus ). As in all nonsauropod sauropodomorphs the ulnar proximal end bears a well-developed olecranon process that extends above the proximal articular surface.
The ulnar shaft is craniocaudally expanded proximally and becomes constricted distally. The cranial margin of the shaft is concave as in Adeopapposaurus , Mussaurus , Aardonyx , Melanorosaurus , and Antetonitrus . The shaft is mediolaterally compressed and triangular in cross-section (with one tip pointing laterally at its proximal third), but becomes elliptical in cross-section along the distal two thirds.
The distal end of the ulna is mediolaterally expand- ed, its maximum dimension being 0.27 times the ulnar length. This condition is similar to that of other basal sauropodomorphs (e.g. Adeopapposaurus , Massospondylus , Aardonyx , Mussaurus , Melanorosaurus , and Lessemsaurus ). More derived taxa, such as eusauropod outgroups (e.g. Vulcanodon and Tazoudasaurus ), however, have the ulnar distal end much less expanded. The long axis of the distal articular surface is parallel to the craniolateral proximal process and perpendicular to the craniomedial process. The distal articular surface is convex.
Radius: Two radii are preserved, with slightly different sizes (BP/1/7135 and 7436). The description is based on BP/1/7435, which presents fewer reconstructed portions with plaster.
The radius is a gracile bone, with the proximal end slightly more expanded than the distal end (maximum mediolateral width of the proximal end is 1.32 times the maximum width of the distal end) ( Fig. 11 View Figure 11 ). The proximal end is subrectangular to suboval, a condition also reported in Plateosaurus , Adeopapposaurus , Massospondylus , Aardonyx , and Melanorosaurus , but unlike the triangular shape present in Mussaurus . The shaft of the radius is rather straight, as in most basal sauropodomorphs, and its cross-section is subcircular. In the proximal half of the radius, over the craniomedial surface and close to the midshaft, there is a noticeable bulge (‘biceps scar’: Yates et al., 2010; fig. 3h, i), also present in Mussaurus , Aardonyx , Melanorosaurus , and Antetonitrus , although in Sefapanosaurus it is much more developed. The distal end of the radius is subovoid, as in other sauropodomorphs. The caudal margin of the distal end has a well-developed, proximodistally orientated ridge and groove, a feature also present in Mussaurus , Aardonyx , Melanorosaurus , Antetonitrus , and Tazoudasaurus . In Aardonyx , however, it is more elongated than in Sefapanosaurus . This structure was regarded as a ‘radial ulnar process’ by Remes (2008) and as a ‘ligament scar’ by Yates et al. (2010). The long axis of the distal articular surface is set at 45° with respect to the long axis of the proximal articular surface, as in most basal sauropodomorphs.
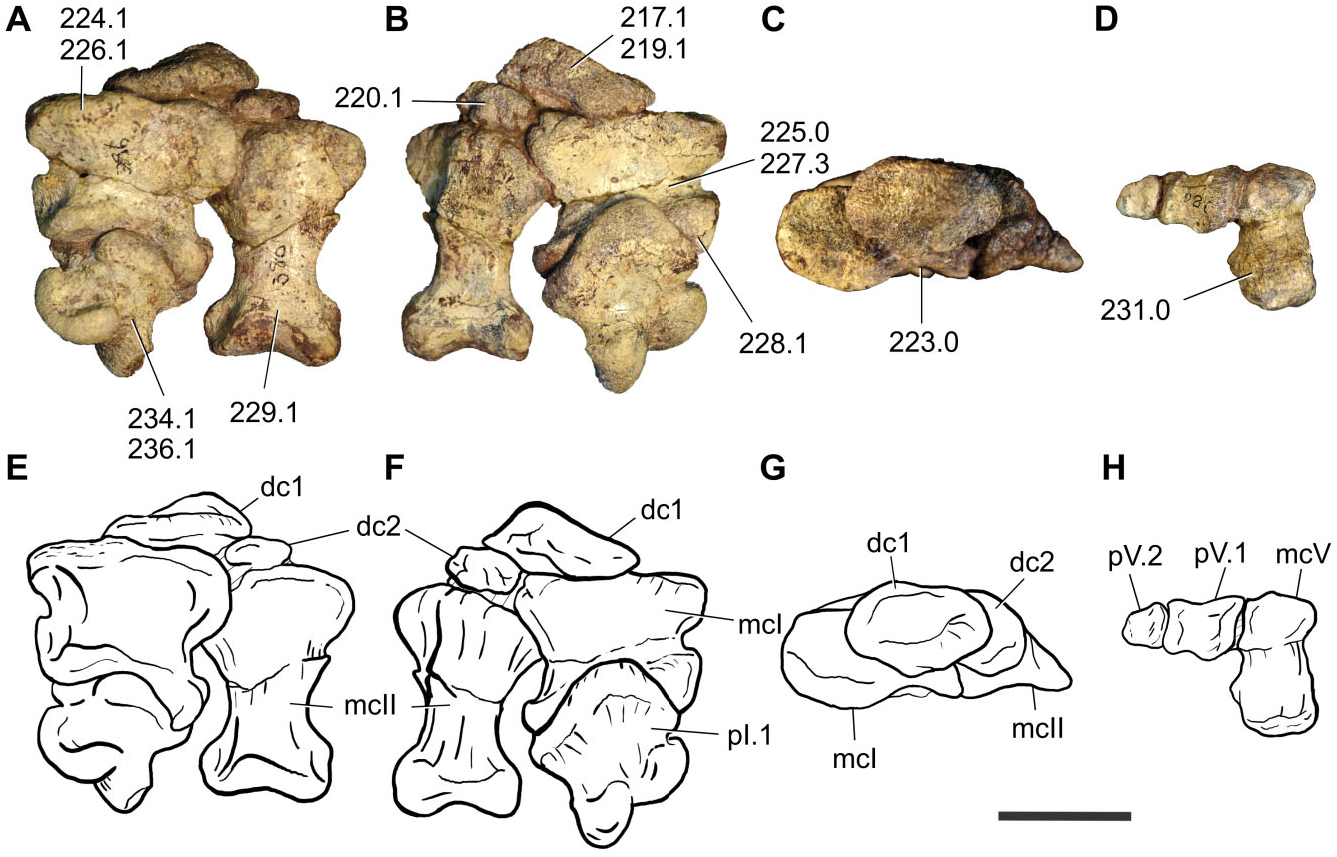
Manus: The preserved elements of the left manus correspond to the distal carpal I and II, metacarpal I, phalanx I.1, and metacarpal II, all of which were preserved in articulation. Additionally, digit five, consisting of a metacarpal, a nonterminal phalanx, and the ungual, is also present amongst the available material (BP/1/7438; Fig. 12 View Figure 12 ).
Distal carpal I is the larger of the two carpal elements preserved. It is suboval in proximal view, with its dorsopalmar length 0.65 times its mediolateral width. Distal carpal I in basal sauropodomorphs is usually the largest of the distal carpals, and varies in shape from suboval (e.g. Adeopapposaurus ) to subcircular (e.g. Massospondylus ), except for Mussaurus in which distal carpal I is notably small (equal in size to distal carpal II) and is subtriangular in proximal view ( Otero & Pol, 2013). Distal carpal I is proximodistally low and subtriangular shaped in dorsal and palmar views. It is mediolaterally narrower than the proximal mediolateral width of metacarpal I, differing from the condition of most basal sauropodomorphs ( Upchurch et al., 2007; Yates et al., 2010) in which distal carpal I covers almost the entire proximal surface of metacarpal I. In this sense, the condition present in Sefapanosaurus resembles that of the most basal sauropodomorphs (e.g. Efraasia , Ruehleia ; Upchurch et al., 2007) and outgroups ( Heterodontosaurus , Herrerasaurus ). As noted above, the basal sauropodiform Mussaurus patagonicus ( Otero & Pol, 2013) also has a small distal carpal I, resembling in this aspect the condition of Sefapanosaurus . As in most basal sauropodomorphs with known carpal elements, distal carpal I of Sefapanosaurus does not present the sulcus across the medial side, and partially overlaps distal carpal II.
Distal carpal II is smaller than distal carpal I, is subcircular in proximal view, and does not completely cover the proximal surface of metacarpal II, as in most basal sauropodomorphs, except for Mussaurus ( MLP 68- II- 27- 1 specimen A), in which distal carpal II is subtriangular in proximal view and the dorsal portion is thicker than the palmar margin.
Metacarpal I is broad and much shorter than the other metacarpals, as in most basal sauropodomorphs. The overall shape of the proximal end is partially obscured by distal carpal I. However, it can be noted that the proximal end is mediolaterally well developed, as in most basal sauropodomorphs. The dorsoplantar height is 0.65 times its mediolateral width, whereas the proximodistal length is 1.2 times the proximodistal length of metacarpal II. The mediolateral width is 0.8 times its proximodistal length, an intermediate ratio between more primitive forms (e.g. Adeopapposaurus , 0.77) and more derived taxa (e.g. Antetonitrus , 1.12, McPhee et al., 2014). The mediolateral length of the proximal end of metacarpal I is 1.5 times the mediolateral width of the proximal end of metacarpal II. A broad proximal end of metacarpal I with respect to metacarpal II is common for non-neosauropod sauropodomorphs (with the exception of Omeisaurus , which has the neosauropod condition). The dorsal and plantar margins of the proximal surface are subparallel to each other and concave, whereas the medial margin is convex and the lateral margin is concave and receives the medial surface of metacarpal II. Metacarpal I is inset into the carpus, as is common for most basal sauropodomorphs except for Sarahsaurus and Thecodontosaurus . The proximal surface is flat and is twisted approximately 45° medially with respect to the distal articular surface.
The shaft of metacarpal I of Sefapanosaurus is reduced, mediolaterally wide, and dorsoplantarly compressed, which is the general shape reported in other transitional sauropodomorphs leading to Sauropoda, such as Aardonyx and the basal sauropods Antetonitrus and Lessemsaurus . The lateral and medial margins are concave, with the latter extremely concave on its ventral margin. A metacarpal I with a notably concave ventromedial margin is present in Lufengosaurus , Aardonyx , Lessemsaurus , and Antetonitrus ; however, the condition present in Sefapanosaurus is much more pronounced than in these taxa. The minimum transverse width of the shaft of metacarpal I is 1.75 times that of the minimum transverse shaft of metacarpal II. This ratio exceeds the value of 2 that characterizes massospondylids and other basal sauropodomorphs ( Riojasaurus , Sarahsaurus, Jingshanosaurus , and Yunnanosaurus ).
The distal articular condyles of metacarpal I are highly asymmetrical, as is common for basal saurischians ( Gauthier, 1986). The lateral condyle is more distally located than the medial condyle; hence, the first manual phalanx of digit I is slightly deflect- ed medially with respect to the rest of the manus. Collateral ligament pits are present and well developed on both distal condyles.
Manual phalanx I.1 is a robust bone with an expanded proximal end and is extremely short so that the shaft is virtually undifferentiated. The proximal articular surface’s mediolateral width is subequal to its dorsoplantar length and is 0.85 times its proximodistal length. Although the proximal end is covered by the distal condyles of Metacarpal I, it can be noticed that it has two distinct articular facets for the articulation of the distal condyles of metacarpal I, as in most basal sauropodomorphs. The ventral margin has an extremely well-developed lip that tapers proximally and covers almost the entire surface of the ventral condyles of metacarpal I when articulated. A well-developed, proximally pointed ventral margin of phalanx I.1 is present in Mussaurus patagonicus ( MLP 68- II- 27- 1 specimen A), although is less developed than in Sefapanosaurus . The distal articular surface is trapezoidal and twisted medially approximately 60° with respect to the proximal end, a condition also present in some basal forms (e.g. Thecodontosaurus, Efrasia ), some ‘core prosauropods’ (e.g. Sarahsaurus , Riojasaurus , Adeopapposaurus ), basal sauropodiforms (e.g. Aardonyx ), and basal sauropods (e.g. Antetonitrus ). The condyles are well developed and separated by a well-defined intercondylar groove. The lateral and medial margins are set obliquely to each other. Collateral ligament pits are well developed and delimited by sharp margins.
Metacarpal II is a robust element with expanded proximal and distal ends. The proximal articular surface is subtriangular with the mediolateral width being 1.2 times the dorsoplantar length and 0.6 times the proximodistal length. The shaft of metacarpal II differs from that of other basal sauropodomorphs in having the lateral and medial margins not parallel to each other. This condition is also present in Lessemsaurus and Antetonitrus . The lateroventral surface presents a bifurcated notch, similar to that reported in Antetonitrus ( McPhee et al., 2014) . The distal articular surface has well-defined condyles, with a slightly developed intercondylar groove, denoting a gynglymoid shape.
Metacarpal V is robust and constitutes the smallest of the preserved elements of the metacarpus, as in most basal sauropodomorphs. The proximal end is almost rounded with its mediolateral width being 0.66 times its proximodistal length. The proximal surface is flat, as in some basalmost sauropodomorphs, such as Thecodontosaurus and Efraasia , but also as in sauropods more derived than Tazoudasaurus . The shaft is short, with a subcircular cross-section. The distal end is subtrapezoidal and lacks differentiation of the distal condyles.
Digit V has a nonterminal phalanx and an ungual. The shape and proportions are similar to those of other basal sauropodomorphs, such as Plateosaurus engelthardti ( MB. R. 4430), Massospondylus carinatus (BP/1/4934), Adeopapposaurus mognai ( PVSJ 610), and Mussaurus patagonicus ( MLP 68- II- 27- 1 specimen A). The nonterminal phalanx has expanded proximal and distal ends. The mediolateral length of the proximal end is 1.5 times the dorsoplantar length, and is similar to the proximodistal length. As in most basal sauropodomorphs in which phalanx V.1 is preserved, the shaft of this element has its lateral and medial margins not parallel to each other, tapering distally. The distal end is differentiated from the shaft but lacks discrete and differentiated condyles.
The ungual phalanx of digit V is also preserved in articulation. This is the smallest element preserved, with an expanded proximal end and a tapering distally rounded end, which lacks differentiated distal condyles.
Hindlimbs
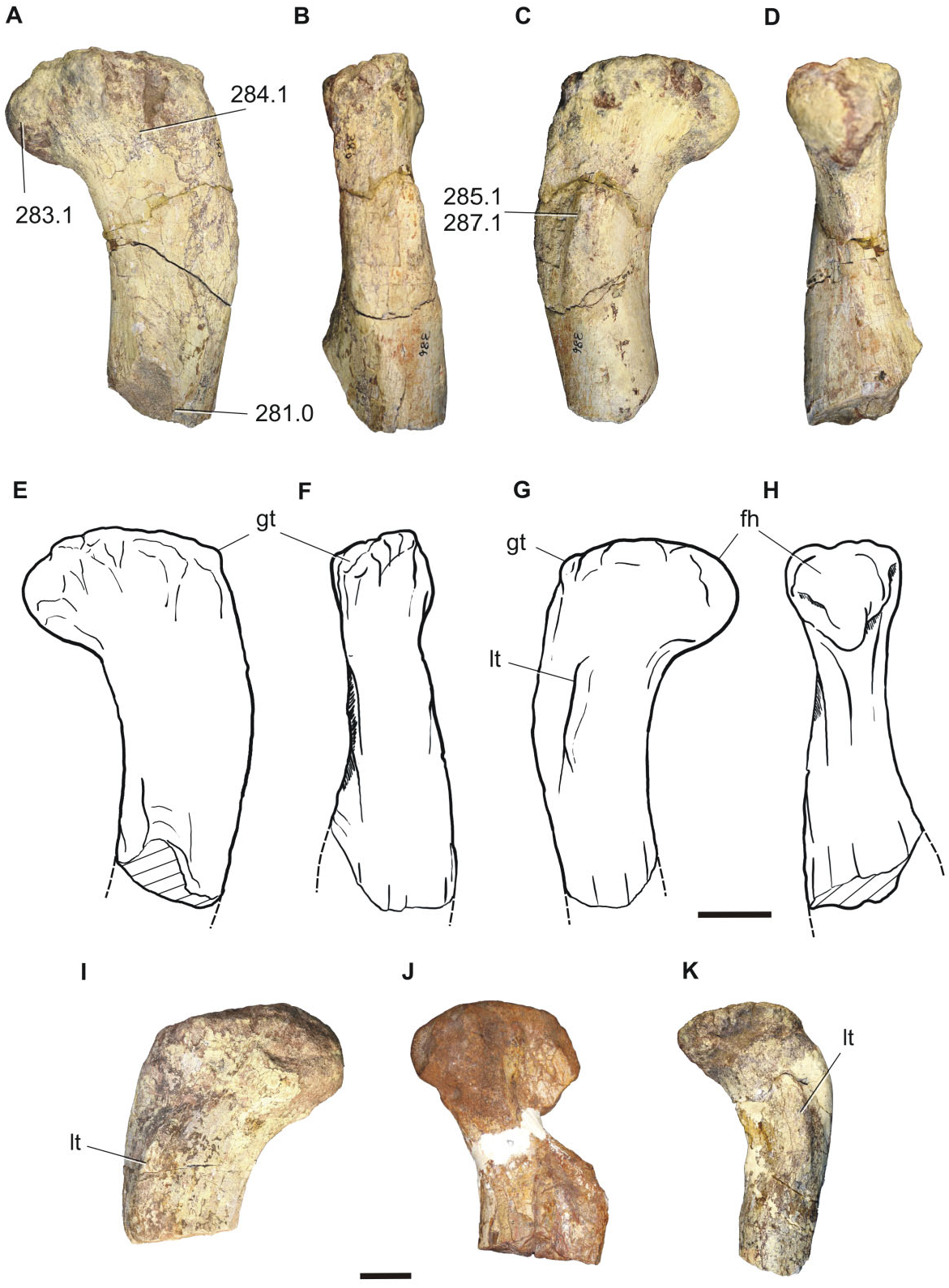
Femur: Four proximal femora are preserved (BP/1/ 7440, 7441, 7442, 7443), which probably correspond to different individuals, all of them of different sizes ( Fig. 13 View Figure 13 ). The proximal region has a well-developed head, the major axis of which is orientated perpendicular to the longitudinal axis of the femoral shaft. The femoral head is roughly hemispherical in profile, having the same condition as in Riojasaurus , Eucnemesaurus , basal sauropods (e.g. Isanosaurus ), and basal eusauropods (e.g. Patagosaurus , Shunosaurus ). The greater trochanter is well developed and is situated at approximately the same level as the femoral head. The lesser trochanter is evident, corresponding to a proximodistally orientated ridge extending onto the cranial surface of the proximal end of the femoral shaft, below the level of the femoral head. A proximodistally elongated lesser trochanter is the generalized condition for most basal sauropodomorphs (except for Saturnalia , which has a rounded tubercle), and basal sauropods (e.g. vulcanodontids). The fourth trochanter is not preserved in any of the specimens.
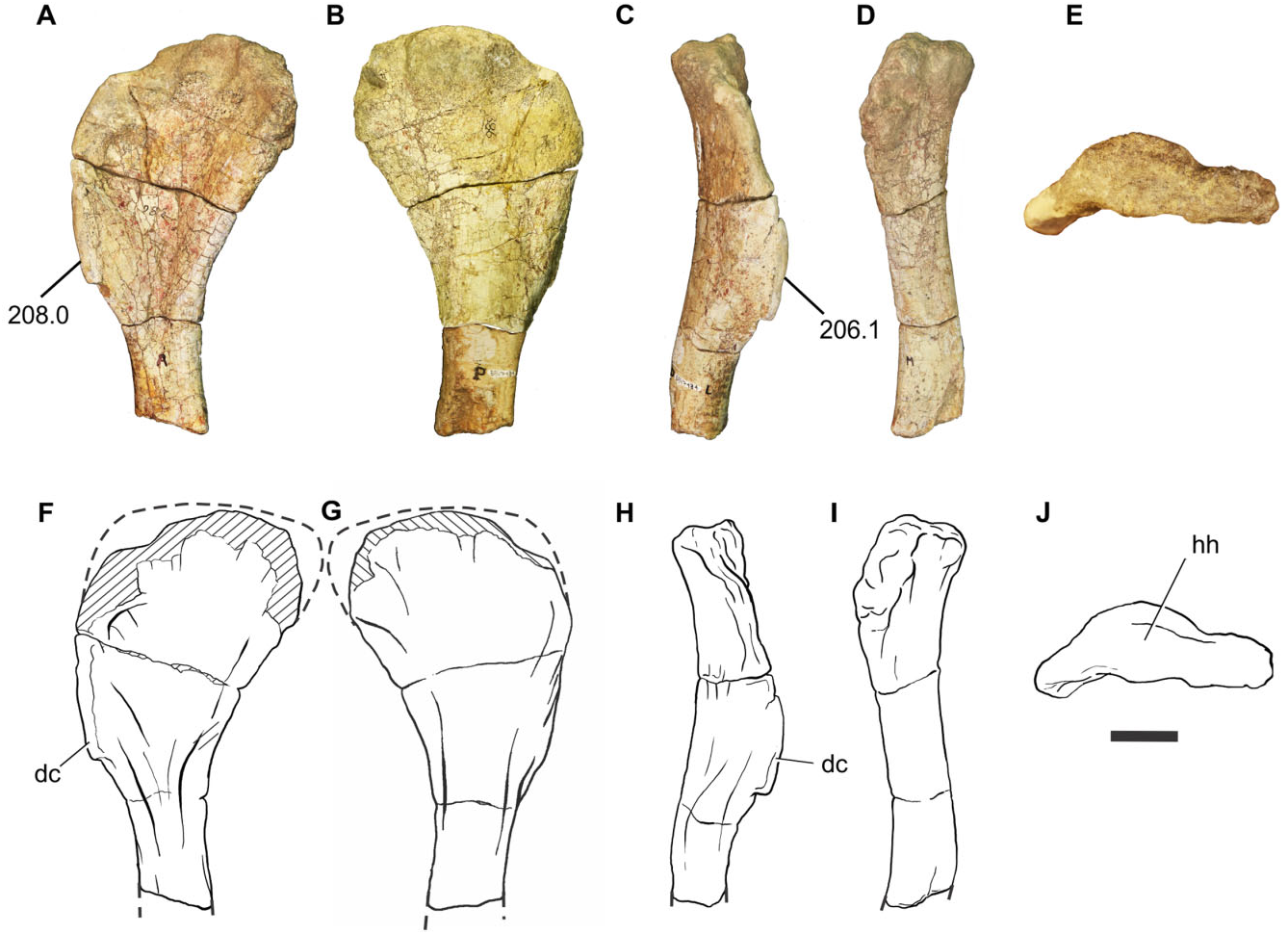
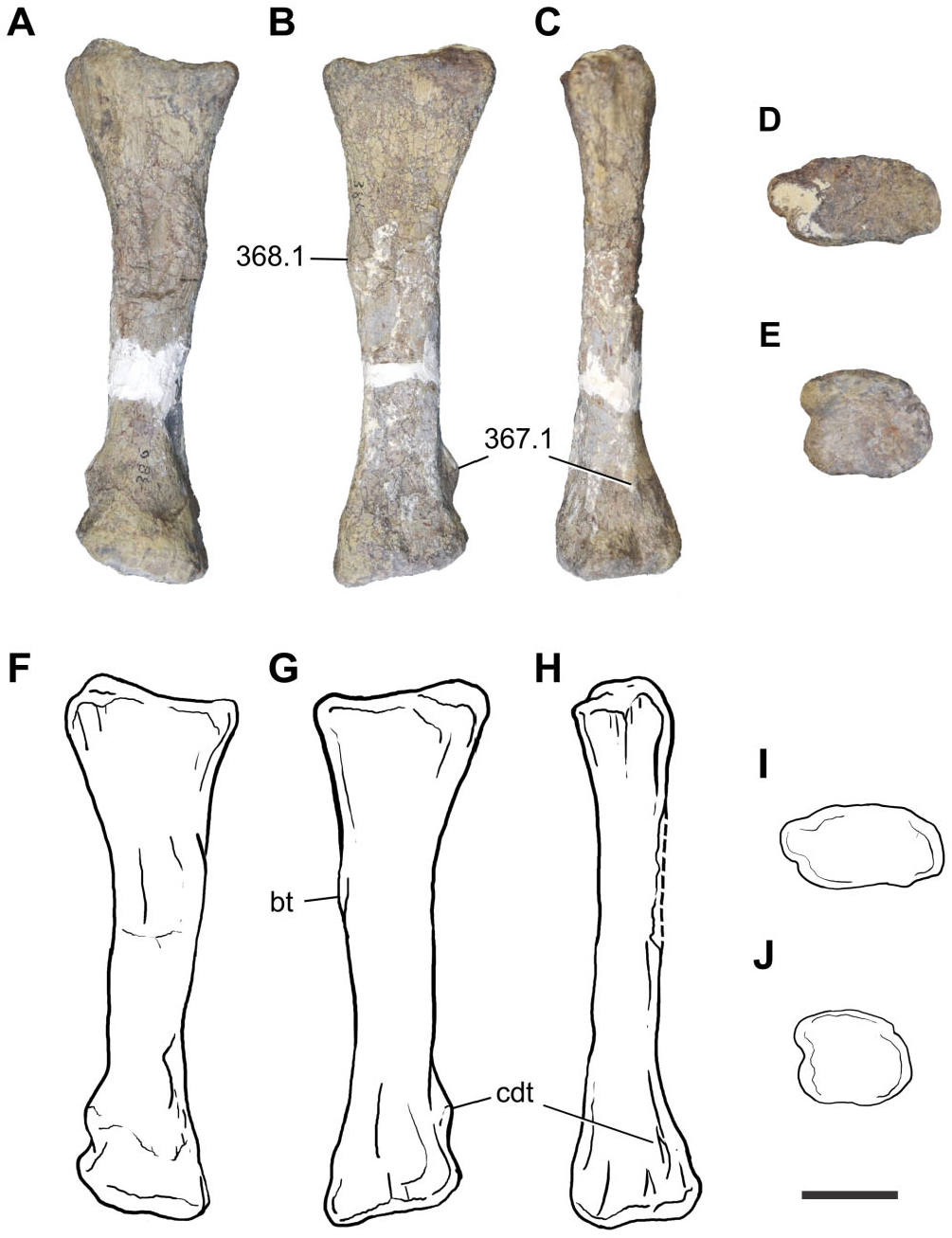
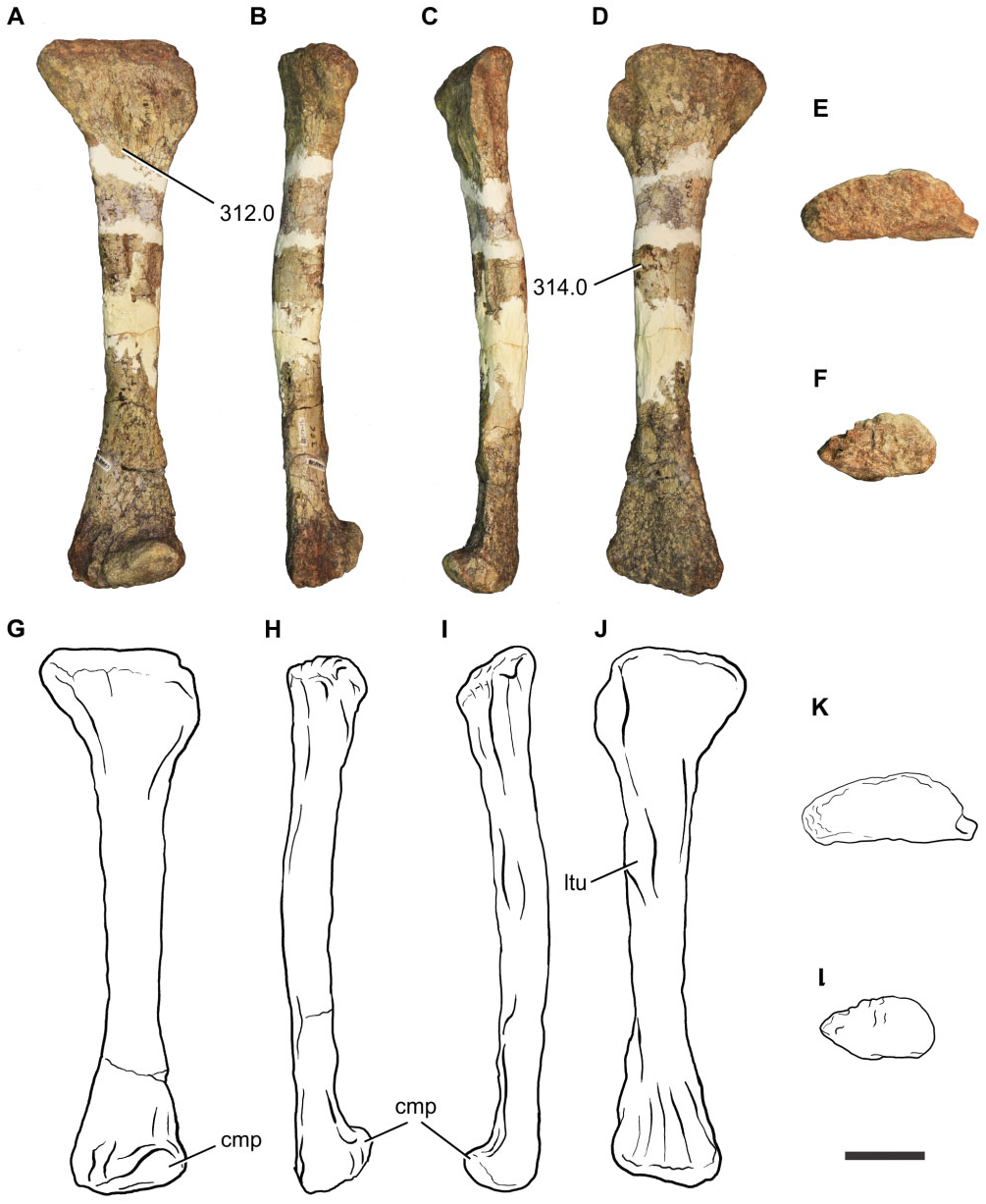
Tibia: A proximal right tibia is preserved (BP/1/7445) ( Fig. 14 View Figure 14 ). The proximal end is well expanded as in all basal sauropodomorphs. The craniocaudal length of the proximal portion is 1.85 times its mediolateral length, giving the proximal end an ovoid shape, with the cnemial crest orientated cranially. The presence of a cranially orientated cnemial crest is characteristic of non-eusauropod sauropodomorphs, whereas the presence of ovoid proximal tibia is a feature present in all non-neosauropod sauropodomorphs (with the exception of Omeisaurus and Mamenchisaurus , which have the derived state; Wilson & Sereno, 1998). The tallest point of the cnemial crest is located close to the proximal end of the crest, as in most nonsauropod sauropodomorphs. The general robustness (maximum craniocaudal length/maximum mediolateral length) of the proximal portion of the tibia of Sefapanosaurus resembles that of Ruehleia ( MB. R.4718.40), Plateosaurus longiceps ( MB. R.4405.57.1, Skelett 1), Mussaurus ( MLP 68- II- 27- 1 specimen A), and Melanorosaurus (NMQR 1551) , but it is not as robust as that of Antetonitrus (BP/1/4952).
Fibula: A complete left fibula is preserved (BP/1/ 7447) ( Fig. 15 View Figure 15 ). The proximal end is craniocaudally expanded, its craniocaudal length being 2.95 times its mediolateral width. The cranial margin of the proximal end is rounded whereas the caudal margin tapers caudally. The lateral surface of the proximal end is convex and the medial surface is flat. The preserved shaft of the fibula is almost straight in lateral view and subcircular in cross-section. The lateral surface of the shaft bears a tenuous lateral tuberosity located close to the proximal end. The distal end is teardropshaped and bears a prominent projection on the craniomedial surface, which represents a unique feature amongst basal sauropodomorphs.
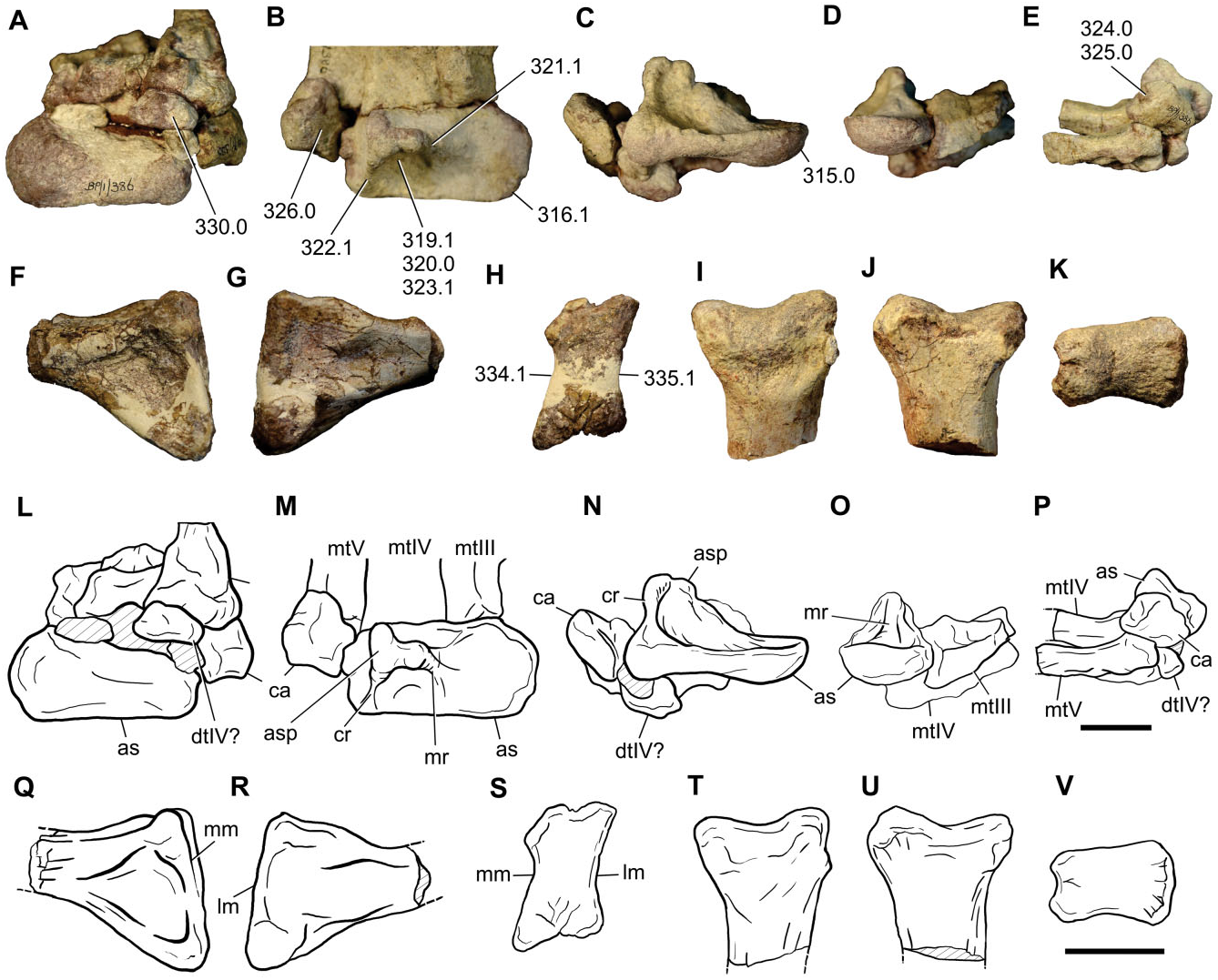
Tarsals and pes: A partially articulated left pes is preserved, including the astragalus, calcaneum, a putative distal tarsal IV, and proximal shafts of metatarsals III, IV, and V, which correspond to the holotype (BP/ 1/386). Additionally, an isolated proximal end of metatarsal II (BP/1/7448) and a distal end of metatarsal III (BP/1/7449) are preserved ( Fig. 16 View Figure 16 ) .
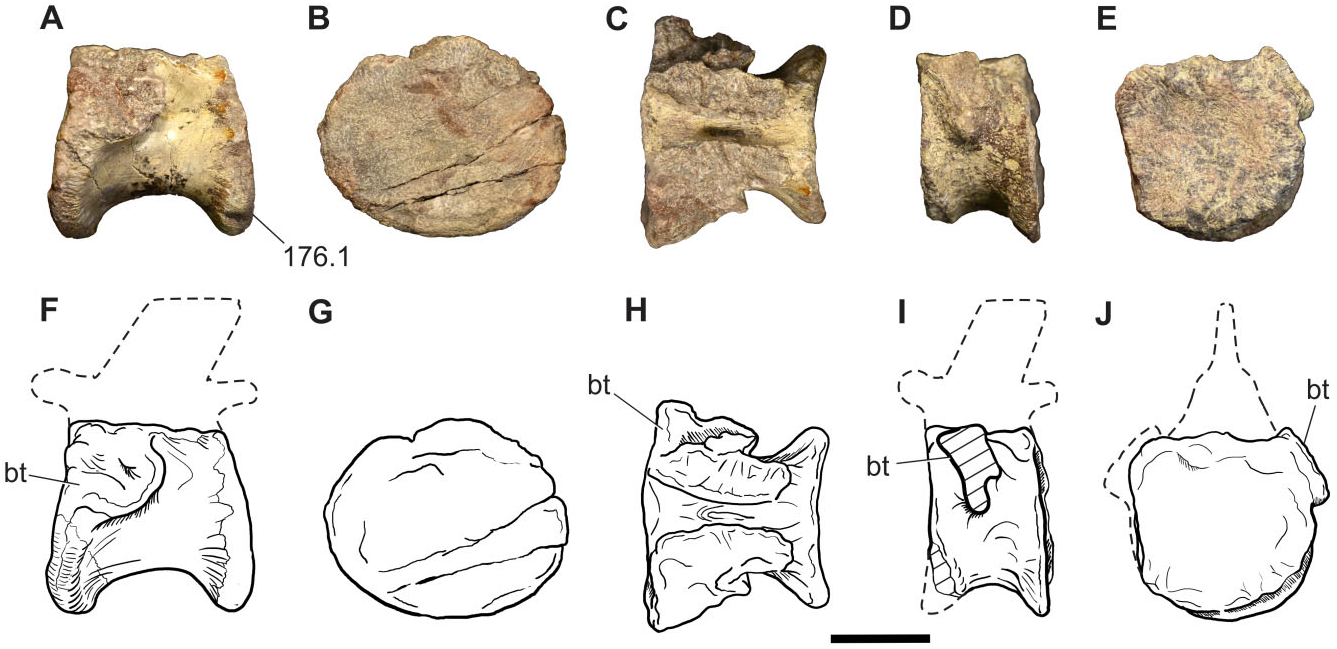
The astragalus of Sefapanosaurus has the generalized condition of most basal sauropodomorphs in being subrectangular in proximal view, with a craniocaudally rounded medial margin and with the lateral surface slightly concave for its articulation with the calcaneum. The lateral and medial margins are almost equally developed craniocaudally, as in Plateosaurus , but unlike other sauropodomorphs (e.g. massospondylids, Blikanasaurus , Mussaurus, Sauropoda ), in which the medial margin is craniocaudally narrower than the lateral margin. In cranial view the astragalus has approximately the same depth both laterally and medially. The caudal margin is concave and lacks the caudomedial convexity reported for Plateosaurus , Mussaurus , and Blikanasaurus ( Otero & Pol, 2013) . The proximal surface of the astragalus is composed by cranial and caudal concave surfaces, which are divided from each other by the ascending process. Both surfaces are approximately equal in size given that the ascending process is placed close to the midpoint of the astragalar proximal surface. Nonetheless, the caudal surface is slightly larger than the cranial one.
The ascending process of Sefapanosaurus has a unique morphology that differentiates it from all other sauropodomorphs. As in all sauropodomorphs, the ascending process is mediolaterally wide and its lateral margin is proximodistally taller than its medial margin. However, in Sefapanosaurus the astragalus is notably tall and its lateral surface is much broader (anteroposteriorly) than the medial surface, and these surfaces are delimited by well-developed ridges that run proximodistally along the entire ascending process. These ridges are unique amongst basal sauropodomorphs because they are notably thick, clearly framing the ascending process, are also straight, and present a subcircular cross-section. This morphology contrasts with the widespread condition amongst basal sauropodomorphs in which the ascending process presents thin framing ridges with convex margins (e.g. Glacialisaurus , Massospondylus , Mussaurus , Blikanasaurus ). The ascending process presents a triradiate ‘T’-shaped cross-section when viewed proximally, representing a unique feature amongst sauropodomorphs, and is regarded here as an autapomorphy of this taxon.
This unusual morphology of the ascending process was probably not caused by injury or disease. Bones affected by traumatic or physiological stress often present characteristic features that are usually preserved in the fossil process (e.g. Gilmore, 1920; Molnar & Farlow, 1990; Farke & O’Connor, 2007; Tanke & Rothschild, 2010). The external surfaces of affected bones are often characterized by a differential texture and/ or bone overgrowth in the surrounding area affected by the disorder, which may allow inference of particular pathologies (see Hanna, 2002 and references therein). Such abnormalities are absent in the astragalus of Sefapanosaurus , which casts doubt on a pathological origin for its unique morphology.
Vascular foramina are absent. The distal surface of the astragalus is craniocaudally convex and mediolaterally concave, as in other basal sauropodomorphs (e.g. Massospondylus , Coloradisaurus , Mussaurus ).
The calcaneum is preserved associated (although not articulated) to the astragalus. It has the generalized proportions of most basal sauropodomorphs, with a transverse width greater than 0.3 times the transverse width of the astragalus ( Yates & Kitching, 2003). The lateral surface is subrectangular, unlike the subtriangular condition present in basal forms such as Herrerasaurus (PVSJ-373), Saturnalia ( Langer, 2003) , and Plateosaurus engelhardti ( GPIT Skelett 1), or the suboval shape present in Blikanasaurus . The lateral surface is convex and lacks the fossa or groove present in other basal sauropodomorphs (e.g. Saturnalia , Anchisaurus , Blikanasaurus , Melanorosaurus ) and sauropods. The proximal surface is concave for the articulation with the distal end of the fibula.
A distal tarsal element is represented by a putative distal tarsal IV, articulated with metatarsal V. Its morphology is mostly obscured by its articulation with other bones, although it can be seen that its mediolateral width is greater than its dorsoplantar depth.
The pes includes partially preserved metatarsals II to V. Metatarsal II only has its proximal end preserved, which has the typical hourglass shape of most basal sauropodomorphs, in which the medial and lateral margins are concave for the articulation with metatarsals I and III, respectively. Conversely, Plateosaurus and Unaysaurus display straight lateral margins. The proximal end is dorsoplantarly expanded, with its maximum mediolateral width 0.67 times the maximum dorsoplantar height.
Metatarsal III to V are preserved associated to the tarsus and to each other; hence, their proximal morphology cannot be observed. Metatarsal III and IV morphology is mostly obscured by the articulation and incompleteness. However, metacarpal IV presents a mediolaterally expanded proximal end, as in most basal sauropodomorphs.
Metatarsal V is almost complete, lacking only its distal end. This element is flat, has an expanded proximal end, and tapers distally, resulting in a triangular shape as in all basal sauropodomorphs. The proximal end is obscured by the presence of a distal tarsal and the calcaneum.


| V |
Royal British Columbia Museum - Herbarium |
| MB |
Universidade de Lisboa, Museu Bocage |
| R |
Departamento de Geologia, Universidad de Chile |
| SMNS |
Staatliches Museum fuer Naturkund Stuttgart |
| BPI |
Bernard Price Institute for Palaeontological Research |
| MLP |
Museo de La Plata |
| PVSJ |
Museo do Ciencias Naturles |
| GPIT |
Institut und Museum fur Geologie und Palaeontologie, Universitat Tuebingen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.