
Myliobatis Cuvier, 1816
|
publication ID |
https://doi.org/10.11646/zootaxa.3860.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:73432BFC-DD6F-4E62-9C41-FCBBEA1A7D29 |
|
DOI |
https://doi.org/10.5281/zenodo.6127318 |
|
persistent identifier |
https://treatment.plazi.org/id/03F55B39-FF2C-CD6B-FF3A-FD95FDB1FC8C |
|
treatment provided by |
Plazi |
|
scientific name |
Myliobatis Cuvier, 1816 |
| status |
|
Genus Myliobatis Cuvier, 1816 View in CoL View at ENA
Myliobatis Cuvier, 1816: 137 View in CoL ( Type species Raja aquila L., by subsequent designation by Bory de Saint-Vincent, 1828)
Myliobatis Geoffroy St. Hilaire, 1817 View in CoL : Pl. 26. ( Type species Myliobatis bovina Geoffroy St. Hilaire, 1817 View in CoL , by subsequent designation—not available, preoccupied by Myliobatis Cuvier, 1816 View in CoL )
Myliobates Schinz, 1822: 234 , 832 ( Type species Raja aquila L.—regarded as a misspelling of Myliobatis Cuvier View in CoL )
Myliobates Agassiz, 1843 : Pl. D (name only, not available; also preoccupied by Myliobates Schinz )
Holorhinus Gill, 1862: 331 ( Type species Rhinoptera vespertilio Girard, 1856 View in CoL , by monotypy)
Species. Myliobatis View in CoL includes 10 valid nominal species (see Table 1). Myliobatis australis Macleay, 1881 View in CoL has long been considered a southern Australian endemic. However, its relationship with the New Zealand endemic M. tenuicaudatus Hector, 1877 View in CoL is poorly known and Last & Stevens (2009) suggested they may be the same species. These two species are morphologically identical and recent molecular barcode (CO1) data showed that a specimen from New Zealand varied by only 0.15–0.31% divergence from Australian specimens, less than the difference between Australian specimens (0.15–0.50%). Thus, M. australis View in CoL is herein considered a junior synonym of M. tenuicaudatus View in CoL .
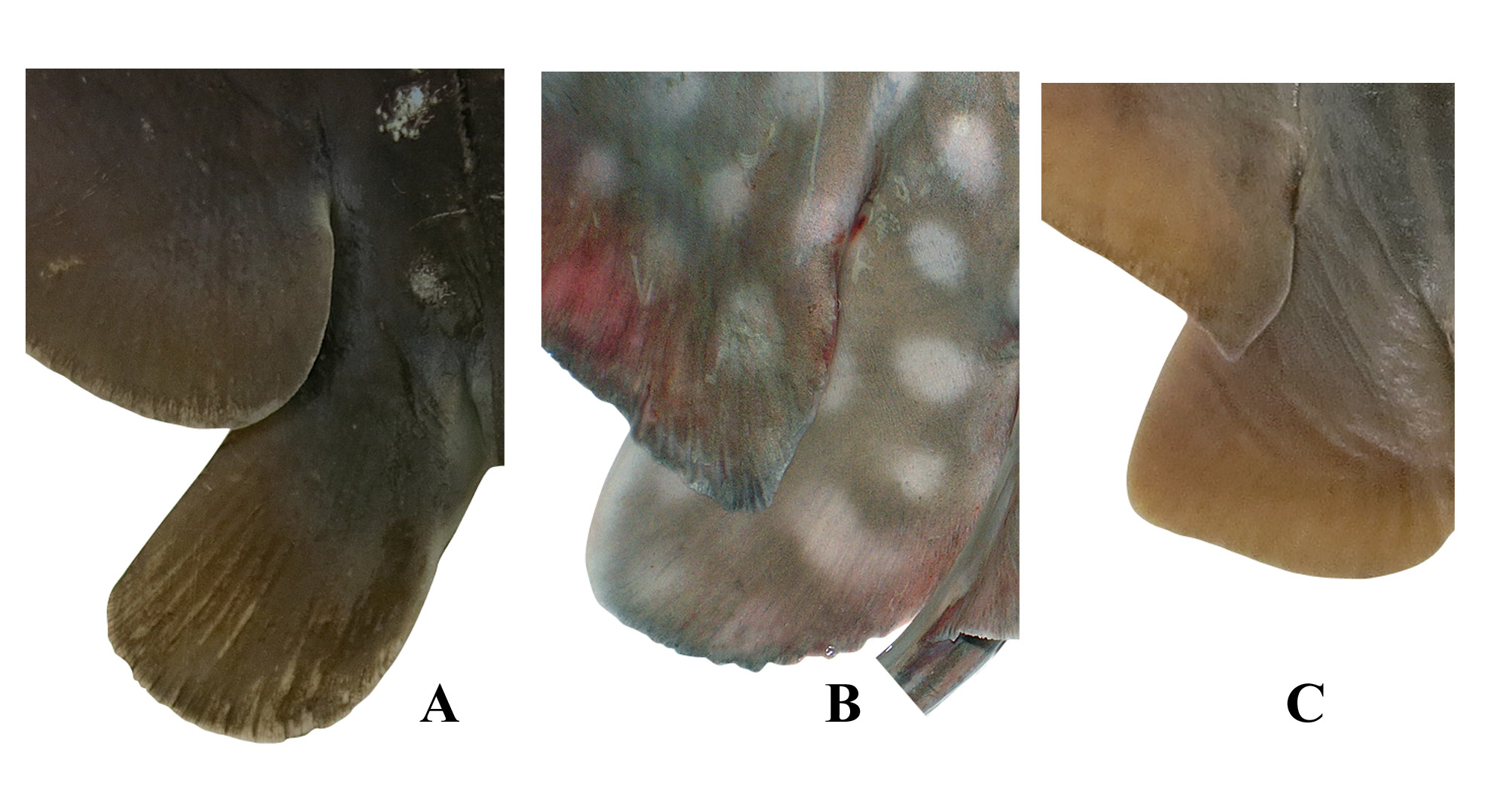
Definition. Myliobatis View in CoL is distinguished from the other two myliobatid genera in the following combination of characters: Anteriormost part of pectoral fins joins head well below eye, joining snout (vs. at level of eye in Aetobatus View in CoL and slightly below eye in Aetomylaeus View in CoL , Fig. 1 View FIGURE 1 ). Rostral part of pectoral fins (snout) connected to pectoral disc by subocular ridges below eyes that are supported by pectoral radials (vs. not connected to pectoral disc in Aetomylaeus View in CoL and Aetobatus View in CoL ); pectoral radials continuous below eyes onto rostral lobe (vs. pectoral radials interrupted from separate cephalic lobe in Aetomylaeus View in CoL and Aetobatus View in CoL ); rostral radial cartilages no less or only slightly less developed as pectoral radials (vs. much less developed in Aetomylaeus View in CoL and Aetobatus View in CoL ). Free rear tip of pectoral-fin with an angular, somewhat pointed apex (vs. broadly rounded apex in Aetobatus View in CoL , Fig. 2 View FIGURE 2 ). Mesopterygium consisting of several components that all articulate with scapulocoracoid (vs. absent or fused with the scapulocoracoid in Aetobatus View in CoL and Aetomylaeus View in CoL ). Head relatively broad (vs. relatively narrow in Aetobatus View in CoL and Aetomylaeus View in CoL ). Postorbital process of chondrocranium in two parts, a small triangular anterior section and an expanded plate-like posterior section (vs. two parts distally fused in Aetobatus View in CoL and Aetomylaeus View in CoL ); lateral margin of process not prolonged and ventrally protruding (vs. prolonged and ventrally protruded, forming a bar-like projection in Aetobatus View in CoL ). Spiracles lateral on head, openings not visible in dorsal view (vs. dorsolateral on head and openings almost completely visible in dorsal view in Aetobatus View in CoL , Fig. 3 View FIGURE 3 ). Nasal curtain straight or slightly undulated (vs. deeply notched in Aetobatus View in CoL , Fig. 4 View FIGURE 4 ). Teeth usually in seven rows in both the upper and lower jaw, some species occasionally found with more than 7 rows (vs. in a single row in Aetobatus View in CoL ); median tooth row widest, transverse (vs. chevron-shaped in Aetobatus View in CoL ). Hyomandibular Accessory Cartilage 1 (HAC-1) small, bar-like and loosely attached to distal end of the hyomandibular cartilage (see Fig. 22A in Nishida, 1990; vs. absent in Aetobatus View in CoL ). Hypo- and basi-hyoid cartilages reduced to a pair of small, bar-like cartilages (vs. absent in Aetobatus View in CoL and Aetomylaeus View in CoL ; see figs 28A vs. 28B in Nishida, 1990). Dorsal fin with a short but obvious free rear tip, inner margin very short (vs. no free rear tip, posterior margin joined to dorsal surface of tail in Aetomylaeus View in CoL , Fig. 5 View FIGURE 5 ); origin just behind or well behind pelvic-fin free rear tips. One or more barbed stinging spines present on dorsal surface of tail behind dorsal fin (vs. spines absent or less well developed in Aetomylaeus View in CoL ). Puboischiadic bar of pelvic girdle weakly to moderately arched and robust (vs. strongly arched and only moderately robust in Aetobatus View in CoL ; see figs 36K vs. 36L in Nishida, 1990). Total vertebrae (excluding synarcual) 108–114 (vs. 80–97 in Aetobatus View in CoL and 80–86 in Aetomylaeus View in CoL ); predorsal diplospondylous vertebrae 37–48 (vs. 13–31 in Aetobatus View in CoL and 5–20 in Aetomylaeus View in CoL ). Pectoral-fin radials (excluding rostral cartilages) 85–92 (vs. 89–116 in Aetobatus View in CoL and 79–92 in Aetomylaeus View in CoL ).
Remarks. The genus Myliobatis was first proposed by Cuvier (1816) for rays with the following characters: head protruding above pectoral fins; pectoral fins wider than other rays, resembling an eagle; jaws with large, flat teeth, like paving tiles, differing in proportions according to species; tails very long, tapering; serrated sting present. Although R. aquila is mentioned in Cuvier’s description, Bory de Saint-Vincent (1828) is recognised as subsequently designating this species as the type for this genus. The complication of R. aquila possibly being designated as the type species for the genus Aetobatus Blainville, 1816 (published one month prior to Cuvier, 1816) is discussed in the Aetobatus nomenclatural discussion above.
Gill (1862) proposed the genus Holorhinus for Rhinoptera vespertilio Girard, 1856 . This species name is subjectively invalid as is secondarily preoccupied by Myliobatis vespertilio Bleeker, 1852 when placed in Myliobatis . It was subsequently replaced by Myliobatis californica Gill, 1865 . The character used to distinguish Holorhinus from Myliobatis by Gill was “transverse entire snout”, which is somewhat confusing since all eagle rays have a smooth snout and not notched like rhinopterids. Gill later states (in 1865) that Holorhinus is “scarcely generically distinct from Myliobatis ”.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Myliobatis Cuvier, 1816
| White, William T. 2014 |
Holorhinus
| Gill 1862: 331 |
Myliobates
| Schinz 1822: 234 |
Myliobatis
| Cuvier 1816: 137 |