
Halocypretta profunda, Angel, Martin V., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3709.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:43F153E2-B0C0-4F05-A126-CB061877AFB6 |
|
DOI |
https://doi.org/10.5281/zenodo.5614911 |
|
persistent identifier |
https://treatment.plazi.org/id/03F5905B-FFCA-FF8A-FF0B-FAAC95D1FB64 |
|
treatment provided by |
Plazi |
|
scientific name |
Halocypretta profunda |
| status |
|
Halocypretta striata ( Müller, 1906) View in CoL
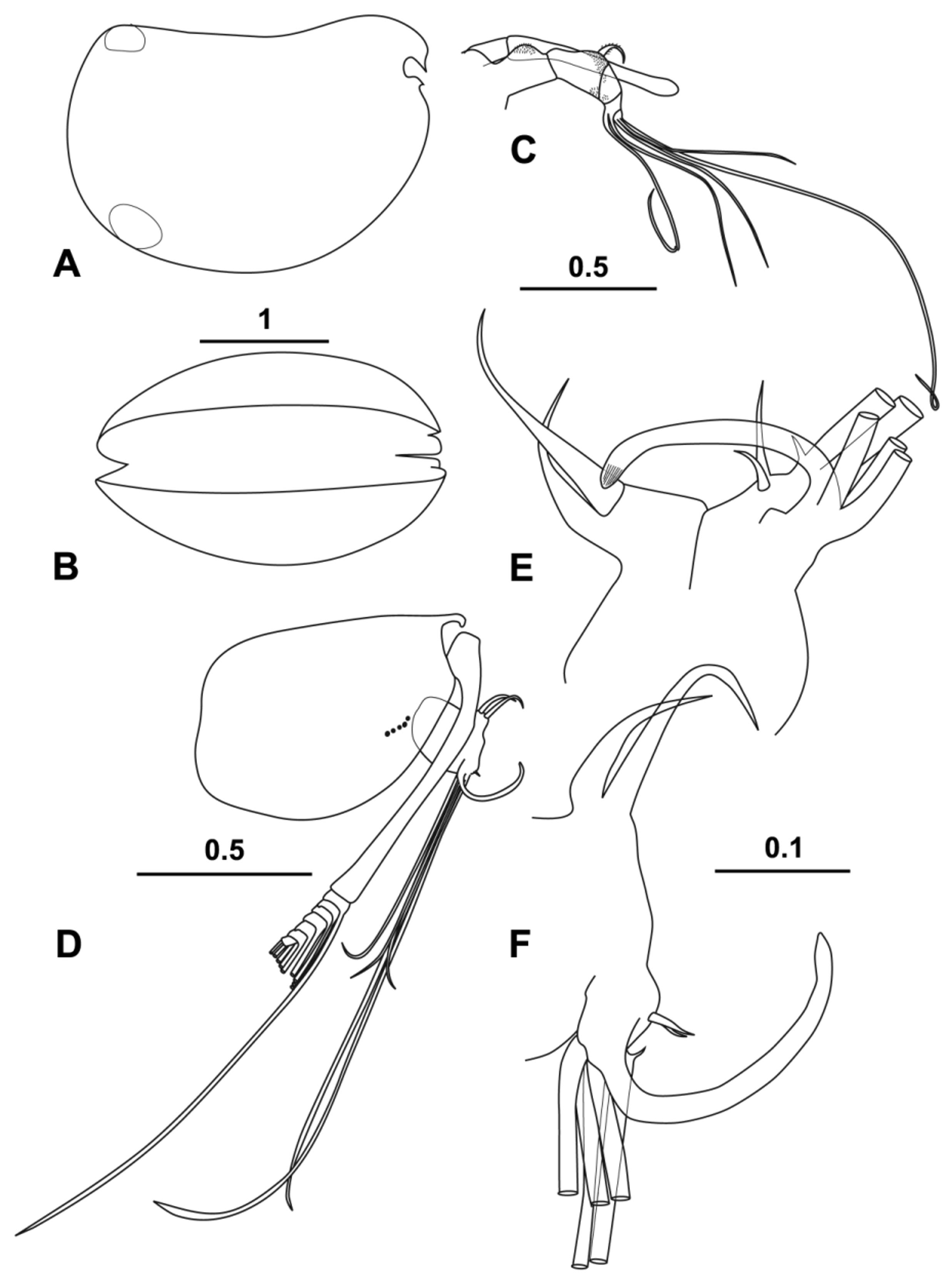
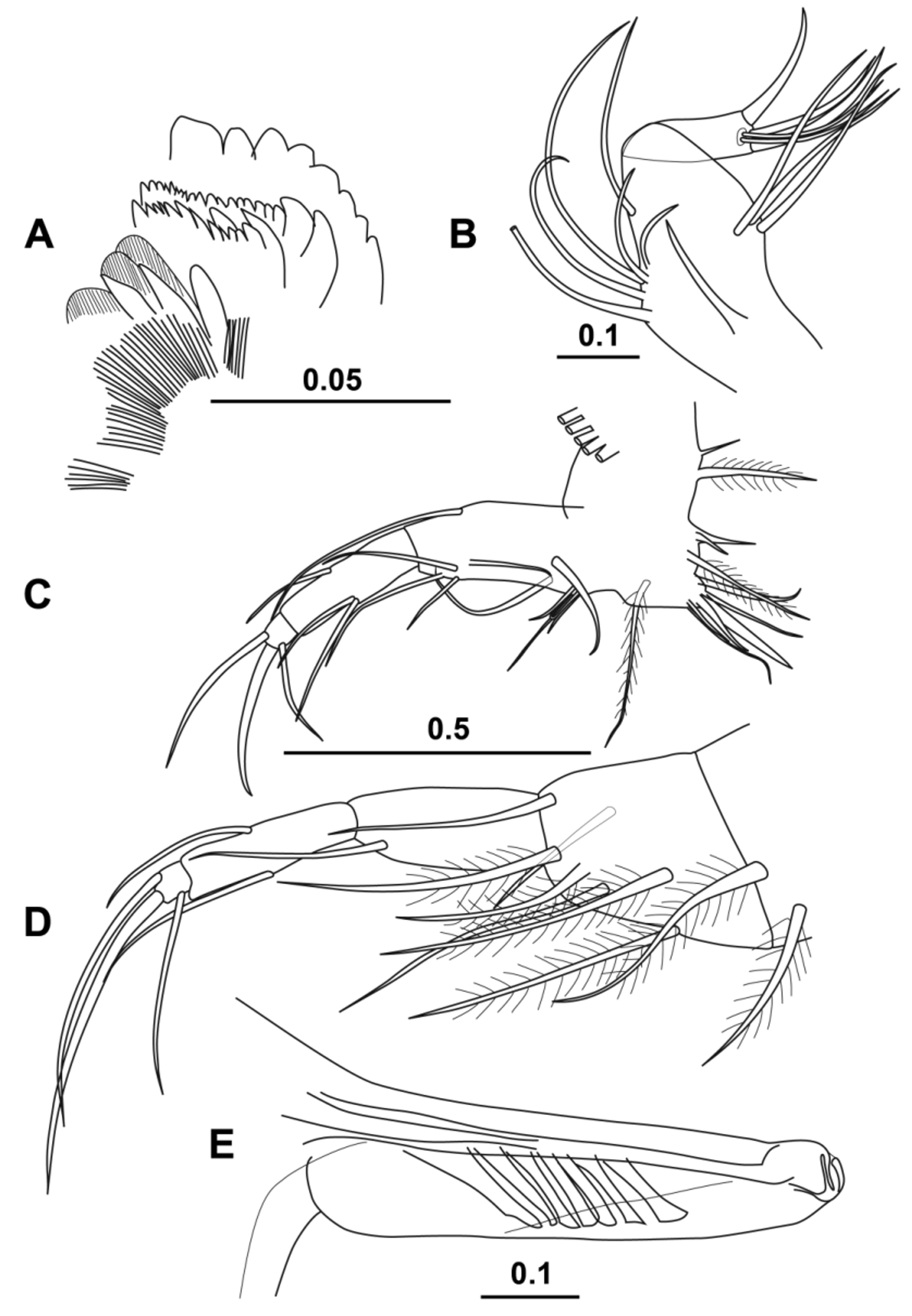
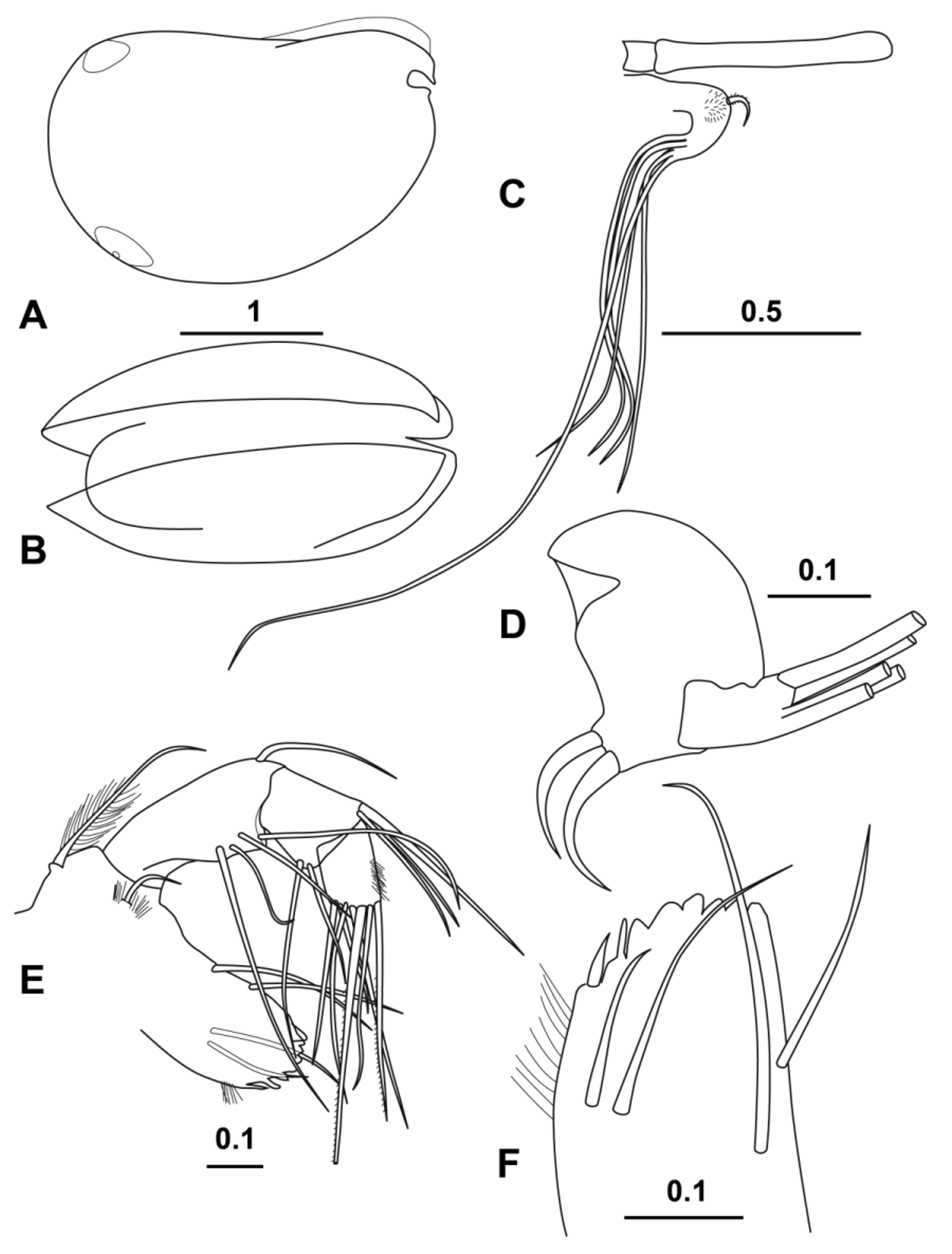
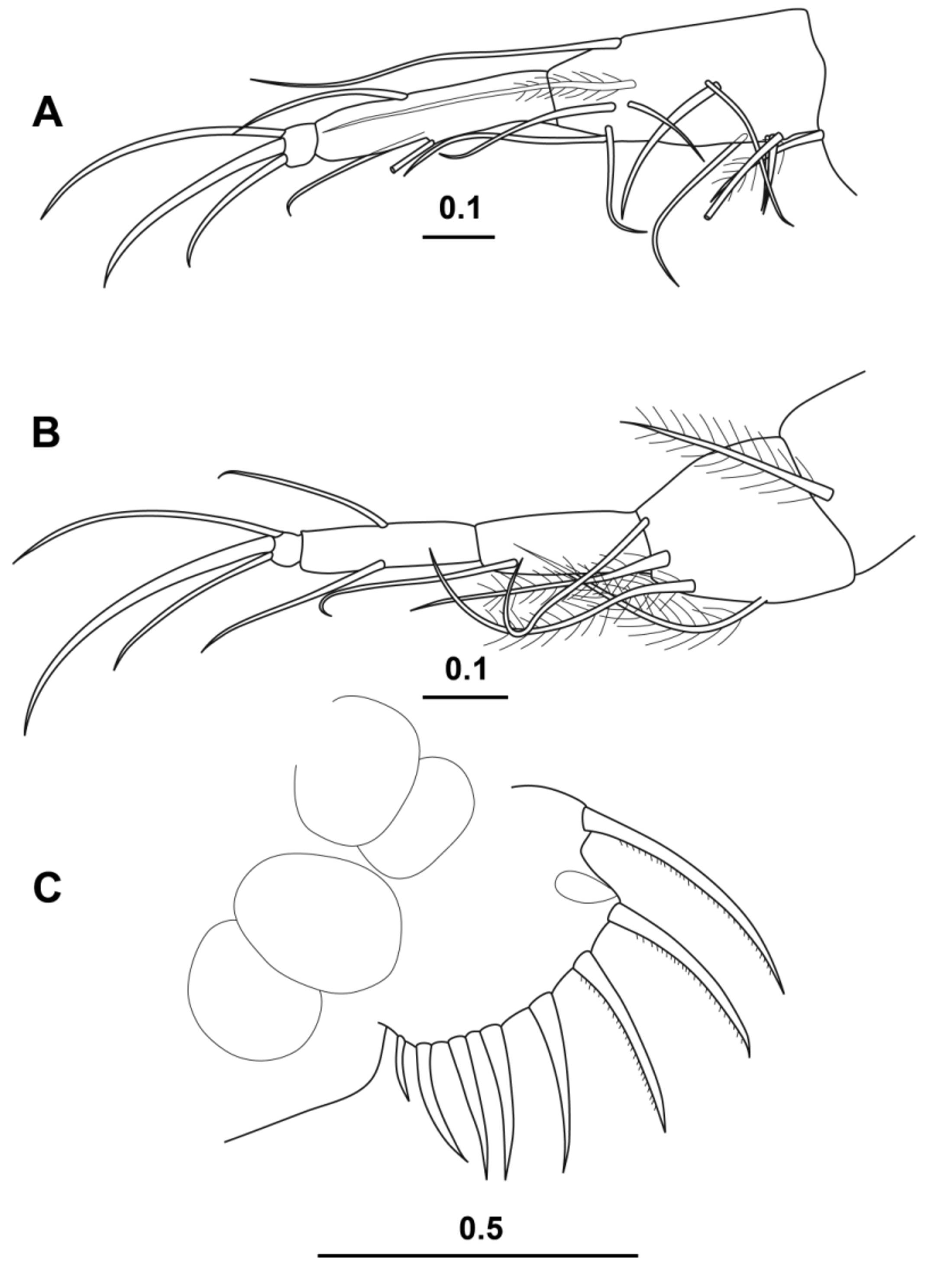
( Figs 13–16 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 )
Müller’s description of a unique male specimen is limited to eleven lines of text and drawings of the carapace, first antenna, frontal organ and hook appendages ( Müller 1906, Tafel VIII (IV), figs 20–23). His male specimen was 2.8 mm long and his figures show the carapace height was 65.5 % CL with a breadth of 72 % CL. His specimen was collected at Tiefsee station 173V in the southern Indian Ocean at 29° 6’S, 89° 39’E in a vertical haul from 2500 m. Since then, there has been only one further tentative record of the species in a sample collected at 37°N 28°W by Granata and Caporiacco (1949). Unfortunately these authors give no description of their specimen, and it would now seem more likely that their record was of Halocypretta profunda . When analysing deep mesoplankton samples from the Gulf of Oman (Charles Darwin Station 54001, hauls 29–31, depths 1387–2009 m, 15 February 1997), Dr Carol Graves found some specimens that she was unable to identify ( Graves 2012). My preliminary examination showed that in all probability they are specimens of Müller’s species. They agree with all the limited information given in Müller’s original description apart from one detail—they lack any carapace sculpture that gave rise to the specific name. However, in many species of halocyprid the expression of striations on the surface of the carapace can be very variable, so there seems no reason at this stage to describe these specimens as a novel species. The specimens were not particularly well preserved, probably because all the specimens would have died while the nets were being retrieved as they passed through the extensive oxygen minimum zone that at the time lay between 150–1250 m ( Graves 2012), and necrosis would probably have set in at least an hour before the samples could be preserved.
Male.
Carapace ( Fig. 13 View FIGURE 13 A, B). The length range of the 17 specimens that were in good enough condition to measure was 2.56–2.88 mm (mean 2.72 ± 0.085) range; carapace height is 58.8 % CL, and breadth is 51.5 % CL, which is in good agreement with the measurements made of Müller’s figure (1906, Tafel VIII (IV), fig. 20). The overall shape is also in good agreement. The rostrum is short (9.7 % CL) and the incisure is very shallow (6.2 % CL). On the lower anterior margin of the incisure is a translucent flange that is a typical feature of the genus, but is somewhat smaller in this species. The opening of the asymmetrical gland on the left carapace valve is level with the posterior end of the carapace hinge, and the one on the right valve is at the posterior ventral corner.
Frontal organ ( Fig. 13 View FIGURE 13 C). As in the other Halocypretta species the stem is very short (5.0 % CL) and the capitulum is long (23.6 % CL) and much longer that the limb of the first antenna. The capitulum is slightly angled ventrally and apart from a barely perceptible curve near its base is straight sided. The distal end is slightly inflated and round. It is bare.
First antenna ( Fig. 13 View FIGURE 13 C). The overall structure is very similar to the other Halocypretta species. The limb is five-segmented and is sharply angled ventrally at the suture between the second and third segments. Its length, not allowing for the angle, is 14.9 % CL. Dorsally at the distal end of the second segment is a short (3.8 % CL) spinose seta that arches anteriorly. Both the first and the second segments have extensive patches of quite coarse bristles. On the first segment there is a patch on the dorsal surface of the segment. On the second segment there is a patch around the base of the dorsal seta and another on the outer flank. The penultimate segment carries two long setae and the terminal segment three. The longest of the terminal setae, the e-seta is 64.9 % CL and is bare. The other four setae are subequal and 35.1 % CL long.
Second antenna ( Fig. 13 View FIGURE 13 D–F). The protopodite is quite short (32.9 % CL). The first exopodite segment is almost the same length (32.6 % CL), and at a quarter length is slightly angled where the muscles flexing the terminal segments are inserted. It also has a small terminal seta. The combined length of remaining segments that carry the swimming setae is 9.2 % CL. The swimming setae are about 42.5 % CL). The shape of first endopodite segment ( Fig. 13 View FIGURE 13 E, F) is roughly oval with a long axis of 9.2 % CL. The a- and b- setae are bare and curved and 4.7 and 7.2 % CL in length respectively. The second segment is quite small. The hook appendages are both C-shaped with pointed tips with slight longitudinal ribbing. There is very little disparity between left and right hooks, which is in agreement with Müller’s (1906) figures (Tafel VIII (IV), figs 22, 23). The second endopodite segment carries short c- and d- setae and a minute e-seta as well as the long f- and g- setae, which are 48.8 % CL and>63.9 % CL respectively. (The g-setae were broken on all the specimens examined). The remaining setae inserted near the bases of the hook appendages are subequal (31.0 % CL).
Mandible. All the mandibles of the three male specimens dissected were damaged, but appeared to be identical to that of the females (see Fig. 15 View FIGURE 15 E). The shape of the limb matches the usual halocyprid form. The first endopodite segment has a bare dorsal seta that barely extends beyond the end of the second segment. There are four ventral setae, one of which is long and inserted on the outer face. The second segment has the usual three terminal dorsal setae, one long and two shorter, and two quite long ventral setae. The terminal segment carries the usual seven setae, of which two are long and claw-like. The longest seta was broken on both sides in all five (male and female) specimens dissected, but is at least half the length of the endopodite.
The toothed edge of the basale has a structure that is typical for most halocyprids with two spine teeth, one sharp, the other blunt, and six cutting teeth. The first of the cutting teeth is offset from the others, and they all have secondary teeth. There is a narrow inner tooth, which is asymmetrically pointed. There are two pairs of lateral setae on the basale and a subterminal seta inserted near the articulation with the endopodite. Around the base of the subterminal seta are patches of quite coarse spines. The exopodite is represented by a single, stout, densely plumose seta.
The toothed edge of the coxale ( Fig. 14 View FIGURE 14 A) consists of eight blunt teeth. The outer tooth list consists of two large teeth and about 16 smaller teeth. The inner toothlist consists of about fourteen sharp teeth of assorted sizes. The masticatory pad consists of a number of clumps of spines and three rather coarse teeth.
Maxilla ( Fig. 14 View FIGURE 14 B). There is a short seta on the basale. The first endopodite segment has three setae on the anterior face of the first segment, and five basal setae and a subterminal set on the posterior face. The terminal segment has three terminal hook setae which are subtended by a pair of long slim setae.
Fifth limb ( Fig. 14 View FIGURE 14 C). The fifth limb has an epipodial formula of 4 + 5 + 4. The basale has ventrally a basal group of four setae and a distal pair, laterally two pairs and a single plumose seta on the outer surface, and a long dorsal seta. The first endopodite segment has a single medial seta dorsally and a pair of medial setae ventrally. The terminal segment carries the usual three setae; the dorsal one being the longest (10.5 % CL).
Sixth limb ( Fig. 14 View FIGURE 14 D). The epipodial formula is 6 + 5 + 5. Two setae are on the coxale; the basale has five setae ventrally, one laterally and a dorsal seta. All these setae, with the exception of the dorsal seta, are plumose. The first endopodite segment carries only a ventral seta at two thirds length. The second segment had a medial seta both ventrally and dorsally. The third segment carries the usual three setae, of which the central one is the longest (13.6 % CL).
Caudal furca. All eight pairs of spines were broken in the three specimens dissected, but overall structure is likely to be similar to the female caudal furca (see Fig. 16 View FIGURE 16 C).
Copulatory appendage ( Fig. 14 View FIGURE 14 E). The appendage is spindle-shaped with a length of 21.4 % CL. The maximum width is 21.3 % of its length. The tip is rounded, and there are six oblique muscles.
Female.
Carapace ( Fig. 15 View FIGURE 15 A, B). The length range of the 18 specimens that were in good enough condition to measure accurately was 2.60–2.96 mm (mean 2.79 ± 0.095 mm). The height is 68.9 % CL and the width 56.2 % CL. The rostrum is 10.2 % CL and the incisures 8.3 % CL. Otherwise all the features are similar to those of the male.
Frontal organ ( Fig. 15 View FIGURE 15 C). The stem is very short (2.1 % CL) and the capitulum is long (22.4 % CL) and bare of any armature. As in the male, the total length of the frontal organ greatly exceeds that of the first antenna.
First antenna ( Fig. 15 View FIGURE 15 C). The length of the limb is 13.9 % CL. The curved, finely spinose, terminal seta on the dorsal side of the second segment is 4.0 % CL. The e-seta, which is flanged distally, is 58.3 % CL and the remaining a–d setae are 27.5 % CL.
Second antenna ( Fig. 15 View FIGURE 15 D). Neither of the two specimens dissected had undamaged second antennae. The length is probably similar to the first exopodite segments (28.5 % CL). As in the male, this segment is slightly angled at a quarter length where muscles flexing the terminal segments are inserted. The combined length of the terminal segments is 9.5 % CL. The longest swimming seta is about 64 % CL, and the shortest terminal seta on the exopodite is 4.2 % CL. The first segment of the endopodite is roughly oval with its longest axis being 7.2 % CL ( Fig. 15 View FIGURE 15 D). The a- and b- setae are bare and curved, and 4.5 and 3.8 % CL respectively. The second endopodite segment is quite short (3.0 % CL). The long setae on the endopodite are bare; f-seta 49.8 % CL, g-seta 71.2 % CL, and the h–j setae are subequal, 30.0 % CL.
Mandible ( Fig. 15 View FIGURE 15 E, F). Is very similar to that of the male. Once again, the longest seta was broken in all the specimens dissected but was>22.4 % CL. The toothed edge of the basale ( Fig. 15 View FIGURE 15 F) is similar to that of the other species.
Maxilla. Similar to that of the male.
Fifth Limb ( Fig. 16 View FIGURE 16 A). The morphology of the limb is very similar to that of the male, and the longest terminal seta (the dorsalmost) is 11.7 % CL.
Sixth limb ( Fig. 16 View FIGURE 16 B). There is little sexual dimorphism shown in this limb. The setae tend to be slightly longer in the male (see Fig. 14 View FIGURE 14 D), but otherwise their numbers and arrangement are similar (some setae on the basale are absent in the Fig. 16 View FIGURE 16 B). However, the longest terminal seta (the central one) is 14.2 % CL), which is marginally longer than the longest terminal seta in the male.
Caudal furca ( Fig. 16 View FIGURE 16 C). In one of the females dissected the caudal furca was intact. There are eight pairs of spines all of which carry fine secondary spines. The decrease in the lengths of the spines along the furca is small except of the final pair. The length of the longest caudal furca spine is 14.9 % CL.
Juveniles.
The carapace lengths of the two juvenile instars identified were: A-1 instar—range 1.72–2.04 mm (mean 1.89 ± 0.105 mm, n = 17) and of the A-2 instars—range 1.26–1.40 mm (mean 1.32 ± 0.039 mm). The bathymetric ranges of the juveniles were the same as for the adults.
Station Date Latitude Longitude Depth (m) Authority Halocypretta parvirostrata
Halocypretta profunda n. sp.
† It seems probable that this record should be referred to H. profunda , but in the absence of any descriptive data or archived specimen this cannot be verified.
* Type locality for each species
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |