
Chavturia abyssopelagica, Angel, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3709.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:43F153E2-B0C0-4F05-A126-CB061877AFB6 |
|
DOI |
https://doi.org/10.5281/zenodo.5614901 |
|
persistent identifier |
https://treatment.plazi.org/id/03F5905B-FFDC-FF98-FF0B-FDF196D8FAD3 |
|
treatment provided by |
Plazi |
|
scientific name |
Chavturia abyssopelagica |
| status |
sp. nov. |
Chavturia abyssopelagica View in CoL , new species
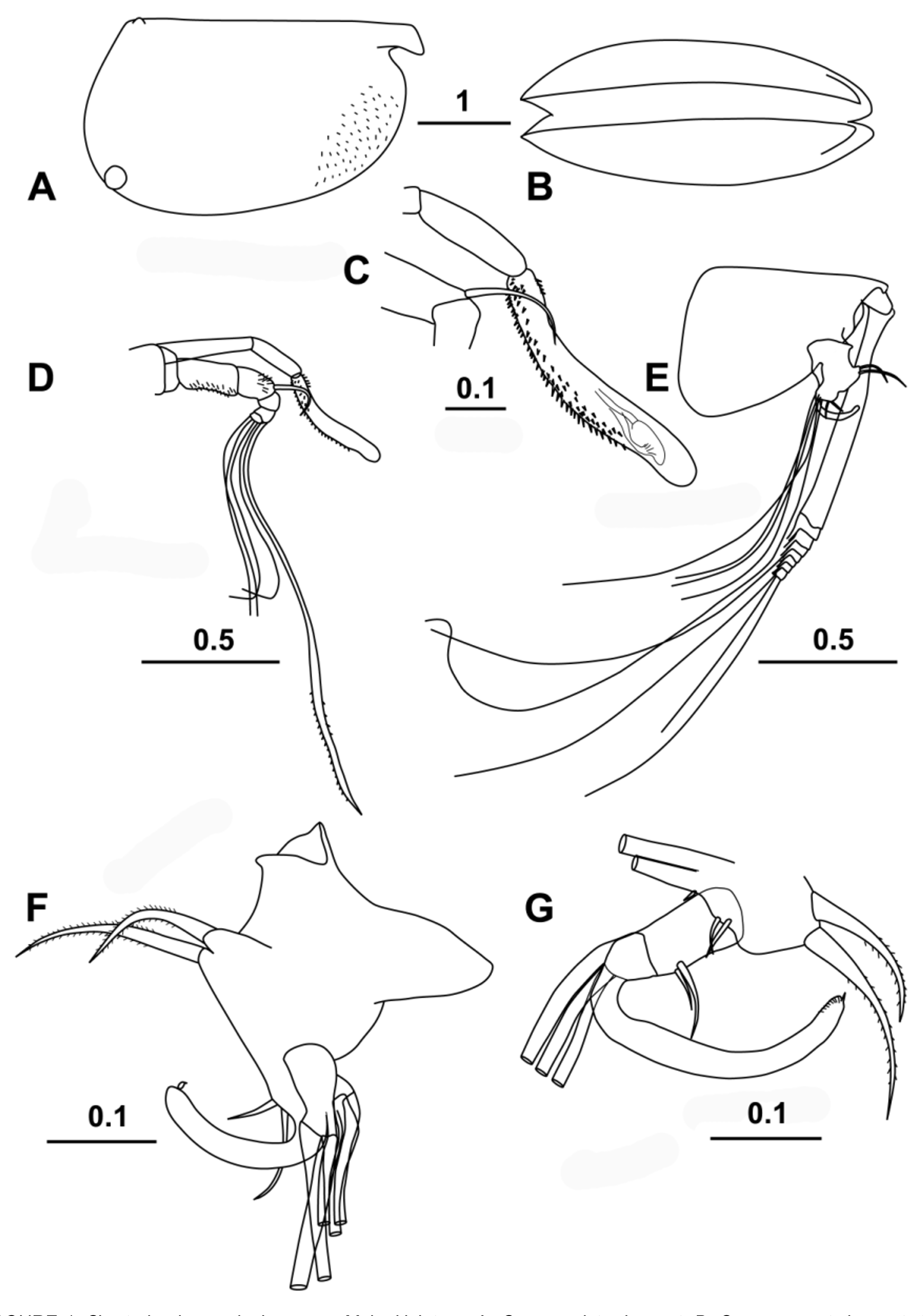
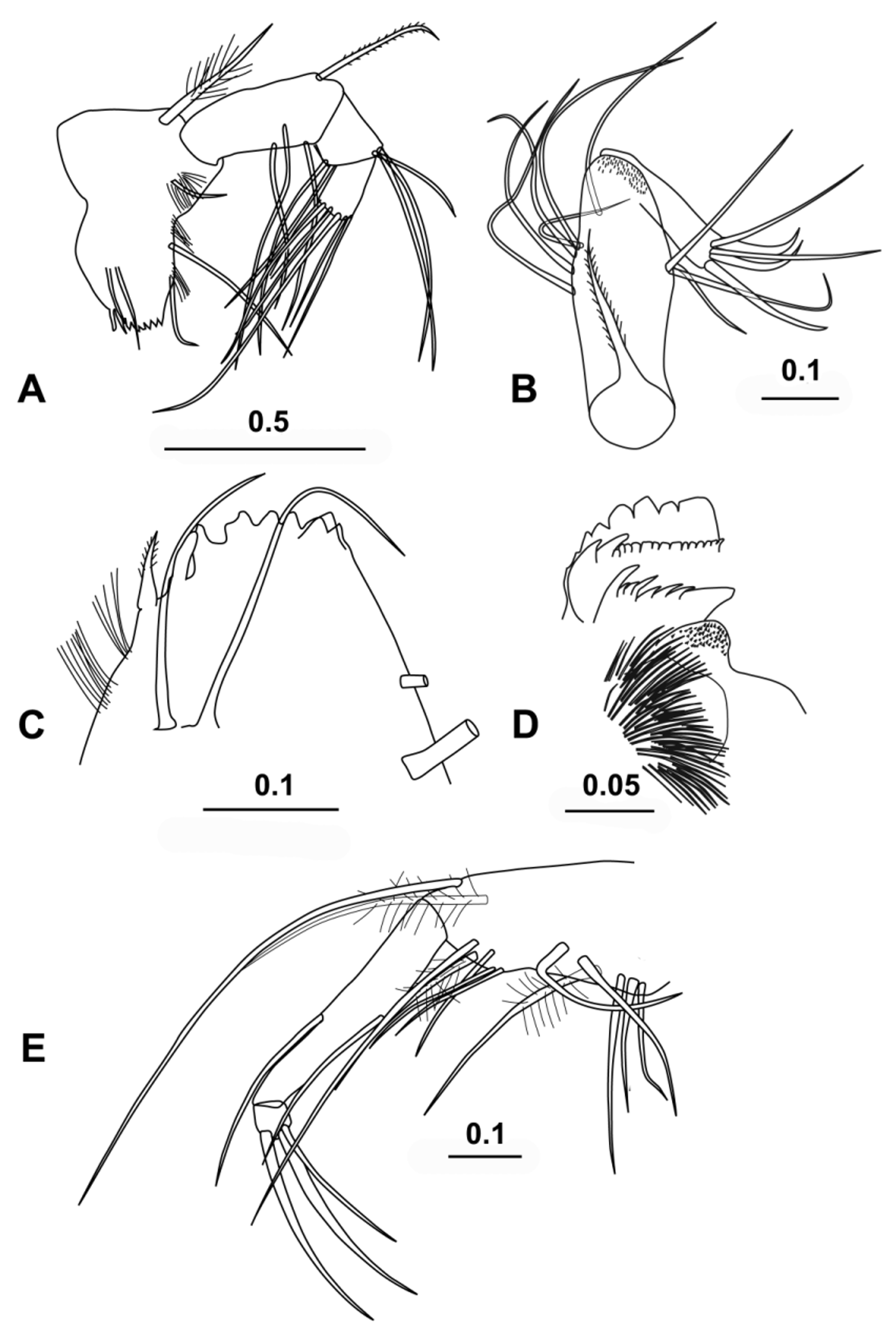
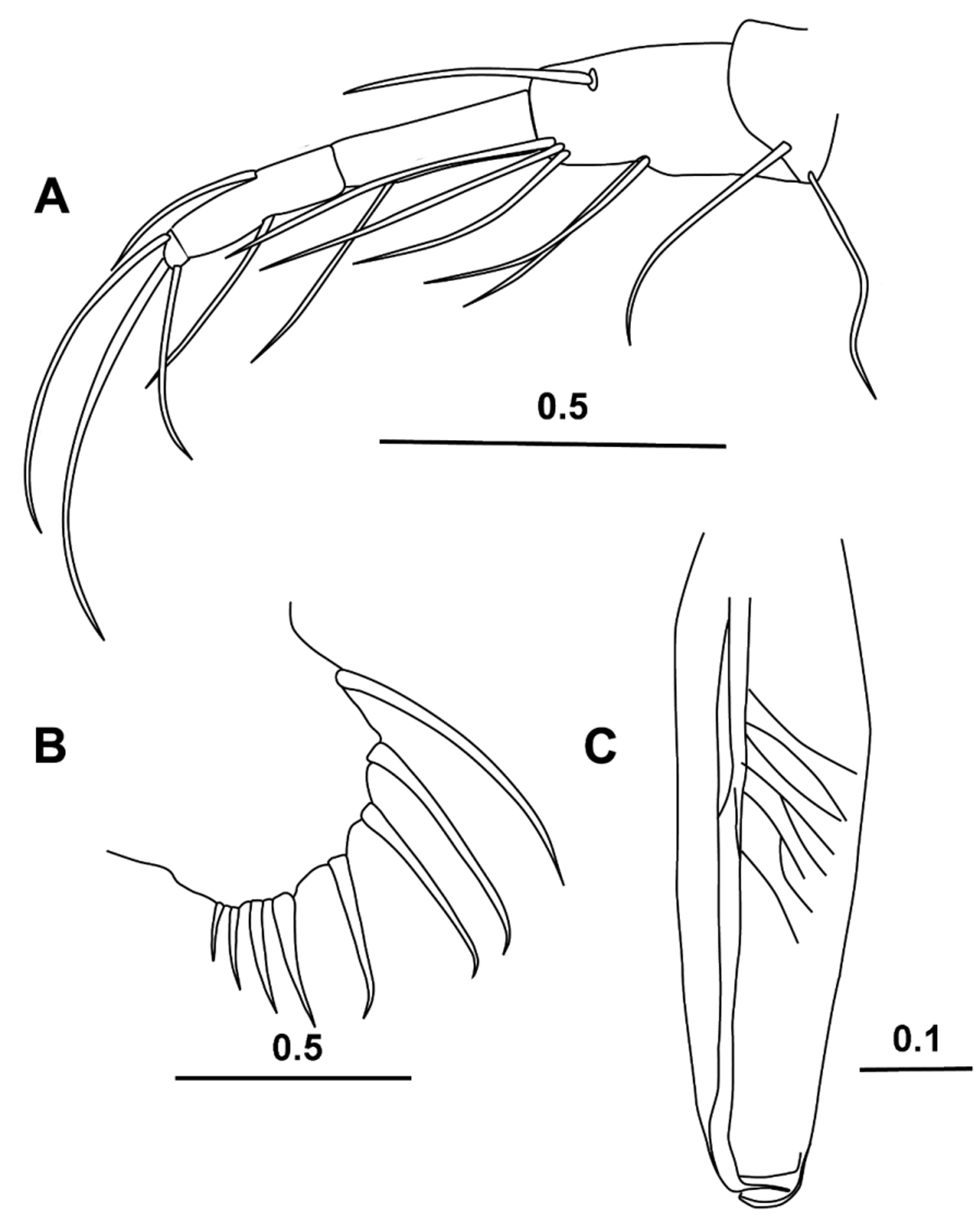
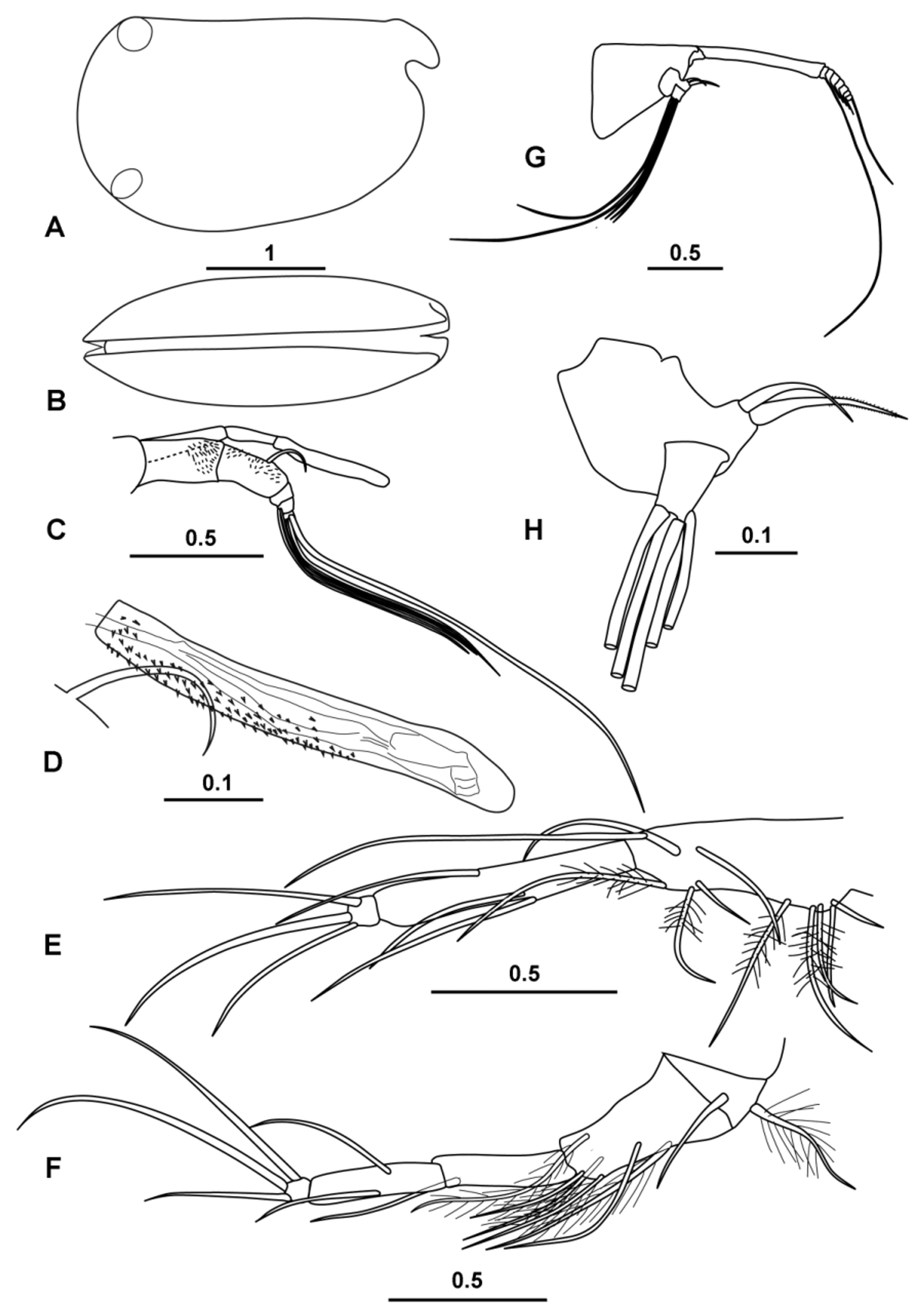
( Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Etymology. The specific name refers to the bathymetric distribution of the species.
Type material. Holotype: 1 male, dissected on slide, NHM registration number 2012.1123 GoogleMaps
Paratypes: 53 specimens, NHM registration numbers 2012.1124 – 2012.1173 GoogleMaps (details see appendix).
Type locality. North-east Atlantic off the North-west African coast at Discovery station 9131#19 GoogleMaps : 20° 14.24’N; 21° 47.06’W; depth 3760– 3510 m; 21 November 1976.
Male.
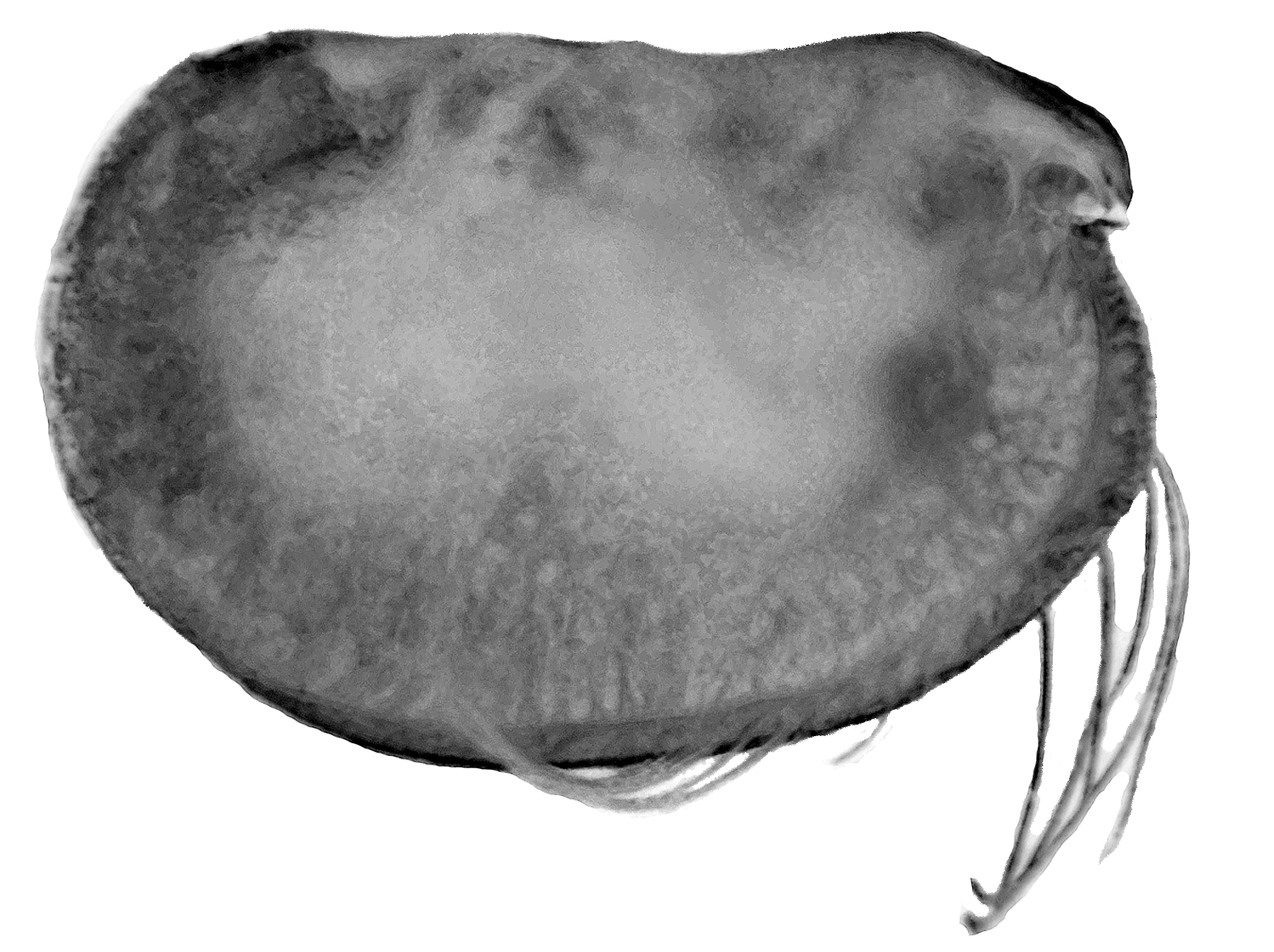
Carapace ( Fig. 1 View FIGURE 1 A, B). Length (CL) ranges 3.16–3.48 mm (mean 3.30 ± 0.104 mm; n = 19) and the height and breadth are 57.5 % and 40.2 % CL respectively (hereafter all percentages of the carapace length are abbreviated to % CL) for the type specimen. The dorsal margin is almost straight. The ventral margin curves smoothly into the posterior margin. The maximum height is at two-thirds the length and there is a marked anterior taper. The rostra are slightly humped dorsally and are quite short; the distance from the anterior end of the hinge between the two carapace valves to the tip of the rostrum is ~10 % CL. The flanks of the carapace are finely spinose in the anterior ventral region below the incisure. The asymmetrical carapace glands are located in the typical positions for the subfamily. On the right carapace valve the gland opens close to the posterior ventral corner (at ~90 % the maximum carapace height below the hinge-line), and directly below the posterior end of the hinge between the two carapace valves. The gland on the left valve opens close to the posterior dorsal corner just anterior (~0.6 % CL) to the posterior end of the hinge between the two carapace valves. In lateral aspect, the incisure is distinct, but quite shallow (7.5 % CL) and is less than the rostral length. In ventral view ( Fig. 1 View FIGURE 1 B) the flanks of the carapace curve smoothly from front to back, and the tips of the rostra are just visible projecting beyond the ventral anterior margin. The live coloration is a uniform red-brown in both sexes and the whole animal is opaque whereas most pelagic halocyprids are translucent ( Fig. 2 View FIGURE 2 ).
Frontal Organ ( Fig. 1 View FIGURE 1 C, D). The shaft of the frontal organ (13.8 % CL) is segmented and the two segments are angled. Even so, the end of the shaft extends just beyond the tip of the limb of the first antenna. The capitulum is a little shorter than the stem (11.9 % CL). It curves anteriorly and its tip is bare and rounded ( Fig. 1 View FIGURE 1 C). On its dorsal surface there are only a few basal spines, whereas laterally and ventrally it is covered with coarse spinules along its basal three-quarters.
First antenna ( Fig. 1 View FIGURE 1 D). The limb is apparently five-segmented, and the last three segments are angled sharply down. The limb’s length (not accounting for the curve) is 14.9 % CL and is only slightly longer than the stem of the frontal organ. The second and third segments are partially covered with coarse setules. Dorsally and terminally the third segment carries a short (4.9 % CL) seta, which arches anteriorly. The fifth segment carries the longest (e) seta (47.8 % CL), which is terminally flanged. The flange is lined with numerous fine spinules along its trailing edge and sparsely along the anterior edge. There are other four setae, two on the penultimate segment and two on the terminal segment, and they are subequal (~22.5 % CL).
Second antenna ( Fig. 1 View FIGURE 1 E–G). The protopodite is relatively short (28.6 % CL), and is only slightly longer than the first exopodite segment (26.6 % CL). The combined length of remaining segments of the exopodite that carry the swimming setae is 6.8% CL, i.e. about quarter the length of the protopodite. The swimming setae are long, nearly 50 % of the carapace length. Consistent with the characteristics of the subfamily, there is no processus mamillaris on the first endopodite segment ( Fig. 1 View FIGURE 1 F). The a- and b- setae are unequal, spinose, curved and similar in length to the first segment. The hook appendage on the endopodite of the right limb ( Fig. 1 View FIGURE 1 G) carries two additional small setae on the inner surface of the hasp as well as the usual three thin walled (h–j) setae that are inserted on the distal end of the hasp. Beyond the hasp the hook angles through ~100°, then straightens and thickens slightly, before it curves again gradually through ~30° to the sub-ridged, rounded end, which is armed with a small thorn. The hook on the left endopodite is a much weaker structure ( Fig. 1 View FIGURE 1 F); it is sickle-shaped with a rounded end that is also armed with a stout thorn. The f- and g-setae are long and simple, 39 % and 59 % CL respectively. The h–j setae are also relatively long, ~24 % CL.
Labrum is slightly curved with about 18 short, stubby flanking filaments.
Mandible ( Fig. 3 View FIGURE 3 A, C, D). The first segment of the endopodite has three long setae on its inner face and a finely spinose subterminal seta on its dorsal face. The second segment has two quite long ventral setae, and three terminal dorsal setae, two of which are long; all these longer setae end about level with most of the terminal setae, the longest terminal seta being the exception. The terminal segment carries seven setae, as is typical for the family. These setae are slim and subequal, apart from the longest seta, which is exceptionally long, 19.4 % CL and 87 % of the limb’s length. The basale has a toothed edge ( Fig. 3 View FIGURE 3 C) with the usual halocyprid compliment of two spine teeth (the first spinose and pointed, the second bluntly rounded and bare) and six triangular cutting teeth; there is also a broad inner triangular tooth that overlaps the last two cutting teeth ( Fig. 3 View FIGURE 3 C). There are two setae inserted on the outer surface, two on the inner edge and one subterminal placed near the base of the exopodite. There are several patches of long setules on the inner surface. The exopodite is represented by a single thick, very plumose seta.
The toothed edge of the coxale has eight triangular teeth ( Fig. 3 View FIGURE 3 D). The outer list has two large curved teeth, one of which is sub-serrate, followed by 14–16 small teeth. The inner list is composed of about 18 uneven teeth.
Maxilla ( Fig. 3 View FIGURE 3 B). There is a long finely spinose seta on the basale. The first segment of the endopodite is elongate and has 6 anterior setae (one offset from the others) and three posterior setae. The end of the segment carries numerous short stubby spines. The terminal segment has the usual compliment of three stout claw-like setae that are subtended by two slim setae, which are longer than the claw setae.
Fifth limb ( Fig. 3 View FIGURE 3 E). The epipodial setae are in three groups: 4 (or 5), 5, 4. The basale has nine setae on its ventral surface arranged in groups of four and five; one seta in each group is plumose. There are two divergent lateral setae, and dorsally there is a longish plumose seta that reaches almost to the end of the first endopodite segment, and a very long subterminal seta that extends well beyond the end of the limb (this is the seta that is considered to be a remnant of the exopodite). The first segment is relatively long and has a single seta both on the ventral and on the dorsal surfaces; both these setae extend beyond the end of the segment. The terminal segment has the usual halocyprid compliment of three terminal setae. The ventral-most of these setae is the shortest and about ¾ the length of the other two, which are subequal (11.3 % CL).
Sixth limb ( Fig. 4 View FIGURE 4 A). The epipodial formula is 7 + 5 + 5. The basale has five ventral setae, a pair at midlength and three sub-terminally. There is no lateral seta but dorsally there is a single seta, which is short relative to the analogous setae in other halocyprid genera, and does not extend to the end of the first segment. The first segment has only a single seta, at ¾’s length on the ventral surface. The second segment has a dorsal and a ventral median seta, both of which extend well beyond the end of the segment. The terminal segment carries three slim curved spines. The central spine is the longest (16.4 % CL) and is double the length of the second segment and just over half the length of the whole limb.
Seventh limb. No data.
Intromittent (copulatory) organ ( Fig. 4 View FIGURE 4 C). The organ is spindle shaped (20.9 % CL) and has four to six oblique muscles and a bluntly rounded end.
Caudal furca ( Fig. 4 View FIGURE 4 B). There are eight pairs of claw setae that are slim and armed with fine spines. The first pair is the longest (20.9 % CL). The first four pairs of spines are curved dorsal-wards near their tips, whereas the smaller dorsal four claw setae curve ventrally. There is no unpaired seta.
Female.
Carapace ( Fig. 5 View FIGURE 5 A, B). The length is 3.43 ± 0.175 mm (n = 19 range is 3.12–3.72 mm). The height is ~58 % CL and the breadth is ~40 % CL and so much slimmer than Halocypretta species. The general features of the carapace are very similar to those of males, apart from the flanks of the carapace being bare of fine spines.
First antenna ( Fig. 5 View FIGURE 5 C). The limb is five-segmented. The first two segments are extensively covered with fine setules, and dorsally the second segment carries a short seta (6.3 % CL) that arches anteriorly. The third segment carries no setae, whereas the fourth carries two, and the terminal fifth segment three. The longest of the terminal setae, the e-seta (46.3 % CL) is bare and is nearly double the lengths of the others (~29 % CL).
Second antenna ( Fig. 5 View FIGURE 5 G, H). The protopodite is short (25.1 % CL) and shorter than the first exopodite segment (26.9 % CL). The endopodite is bare ( Fig. 5 View FIGURE 5 H). Both the a- and the b- setae are curved. The a-seta is bare and about as long as the width of the first endopodite segment. The b-seta is slightly longer and armed with very fine spinules. The terminal segment of the endopodite is cuboid, and it carries five setae as is typical for the family. The bare gseta is the longest (53.4 % CL), and the f-seta is also long (~44.2 % CL) and bare. The remaining three setae, the h-, i-, and j- setae are subequal (~27 % CL), bare and lack shafts.
Labrum. Smooth as in the male, but there are more (25) flanking filaments in the specimen dissected. These filaments are arranged in a double row and so some can be obscured in preparations.
Mandibles and maxillae are similar to those of the male.
Fifth limb ( Fig. 5 View FIGURE 5 E). Only minor differences were noted between the male and female limb. There are eight ventral setae on the basale arranged in groups of five, two and one: a difference may be an artefact resulting from the dissection and orientation of the mounted limb. Laterally, there are two divergent lateral setae, but the dorsal plumose seta appears to be missing, which is probably a dissection artefact. The endopodite segments are very similar, but the terminal three setae are slightly different in length—the dorsal seta is just the longest (11.5 % CL), whereas in the male the central seta that is the longest.
Sixth limb ( Fig. 5 View FIGURE 5 F). There is little sexual dimorphism in this limb, and what differences were noted were relatively minor, for example all the ventral setae on the basale are plumose and the long dorsal seta was missing. The ratios between the lengths of the three terminal setae were similar but the longest seta was relatively slightly shorter (14.9 % CL).
Caudal furca. The female furca is very similar to that of the male, although the longest claw is a little longer (19.4 % CL).
Carapace lengths. The lengths of the 82 specimens collected, 47 adults and 25 juveniles, are summarised in Table 1 View TABLE 1 .
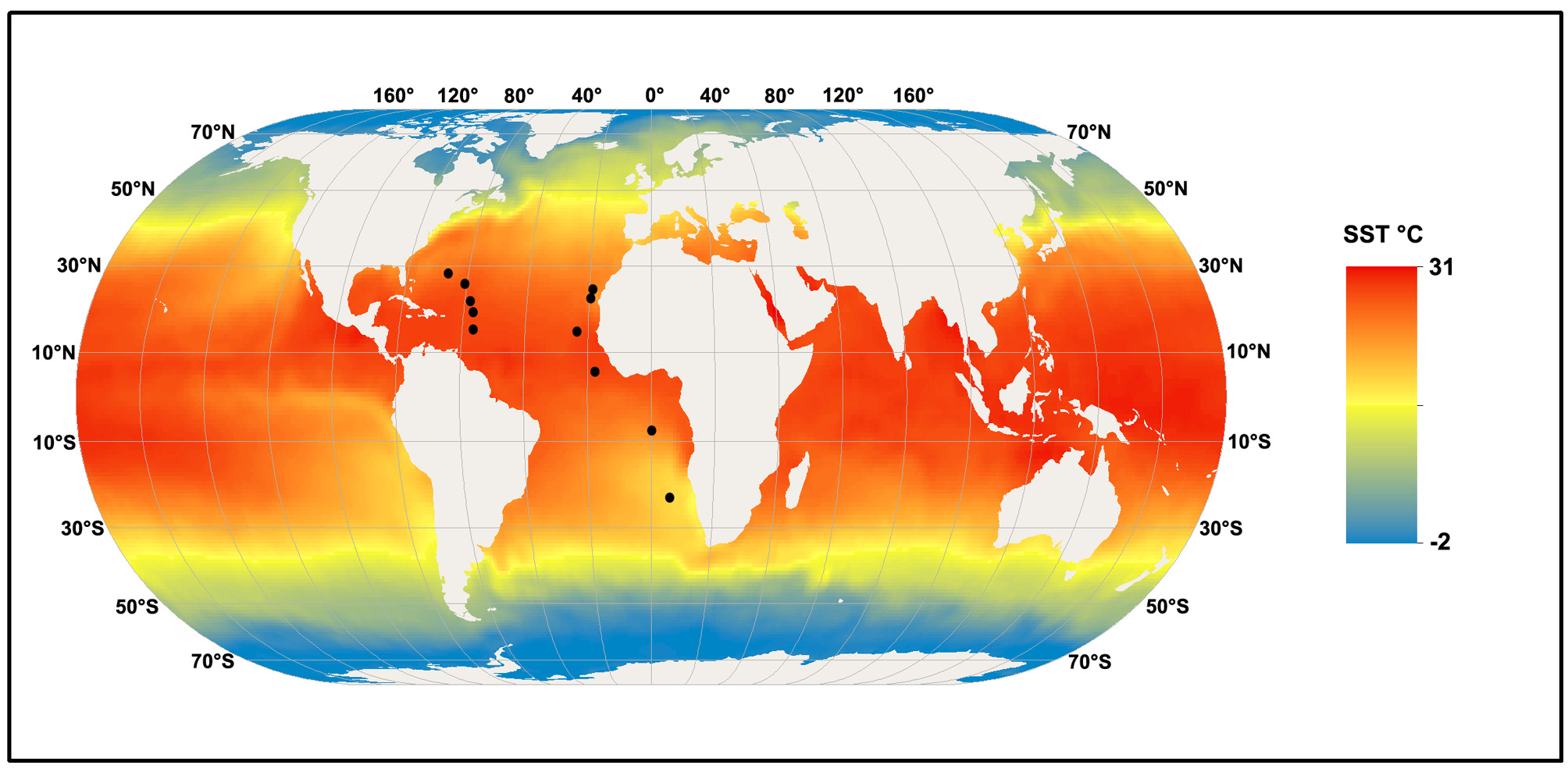
Zoogeographic and bathymetric distribution. The species has a widespread distribution at tropical and subtropical latitudes in the Atlantic ( Fig. 6 View FIGURE 6 ). Within this latitudinal range it has been quite consistently collected whenever sampling for mesoplankton has been carried out at depths> 3000 m. However, there has not been enough deep sampling at latitudes>30° to state categorically that its range is restricted to low latitudes. Nor can it be considered to be endemic to the Atlantic, although presently its known range does appear restricted to the Atlantic. However, its distribution does appear to be related to North Atlantic Deep Water, if so, then it may well be dispersed within this water mass into other oceans.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Halocypridina |
|
Family |
|
|
SubFamily |
Halocypridinae |
|
Genus |