
Mortoniella (Nanotrichia), 2017
|
publication ID |
https://doi.org/ 10.5281/zenodo.5170203 |
|
publication LSID |
lsid:zoobank.org:pub:AB1A57F0-7CB4-4830-920B-DF219740A596 |
|
DOI |
https://doi.org/10.5281/zenodo.5186335 |
|
persistent identifier |
https://treatment.plazi.org/id/03F687A7-FFA7-F867-FF01-BC46412BF8CF |
|
treatment provided by |
Felipe |
|
scientific name |
Mortoniella (Nanotrichia) |
| status |
subgen. nov. |
Mortoniella (Nanotrichia) , new subgenus
Type species: Mexitrichia pacuara Flint, 1974 .
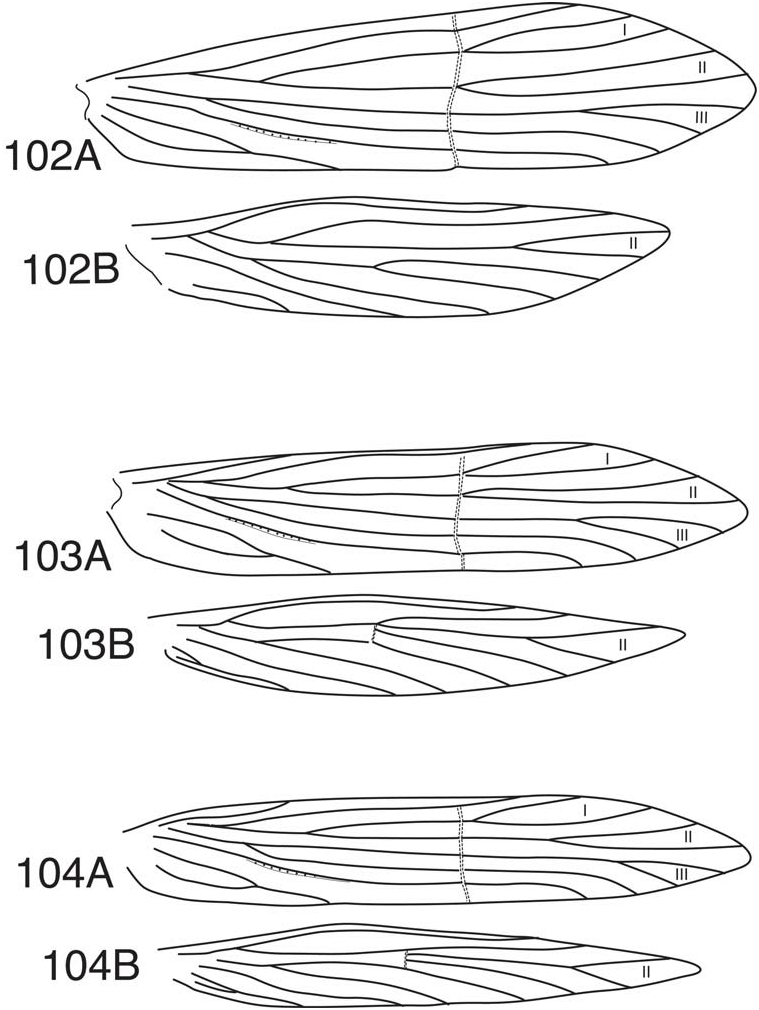
This new subgenus can be distinguished from the subgenus Mortoniella by several characters considered collectively. One of these, the small size of the species, is suggested by the name assigned to the subgenus. The most distinctive character defining the subgenus is the reduction of the hind wing venation to include only fork II. A similar reduction occurs in some species and species subgroups of the leroda group of the subgenus Mortoniella , but generally the costal margin of these species has a distinct bend or inflection ( Fig. 101B). This is much less distinct in species of Nanotrichia , such that the overall wing shape is more spear-like ( Fig. 102B, 103B, 104B View Figures 102-104 ). Another character synapomorphy that seems to be consistent for the included species is that the middle legs have only 3 spurs (rarely 2). A similar reduction occurs in various species in the leroda and bilineata groups of the subgenus Mortoniella , but the usual spur formula in both of these groups is 0:4:4. Additionally, the anterior margin of segment IX of males in the subgenus Nanotrichia is distinctly produced in its ventral ½ and the posterior margin is nearly linear. The lobes of the segment are separated dorsally by ½ or more of the width of the segment (with several exceptions). Species in the bilineata group of the subgenus Mortoniella also have the anterior margin of segment IX produced in the ventral ½ and the lobes widely separated dorsally, but these species are larger and have retained fork V in the hind wing, and usually also fork III. In general, the very shortened phallobase and relatively simple structure of tergum X of species in Nanotrichia also support their separation from the subgenus Mortoniella . Two very distinctive species groups are assigned to Nanotrichia , the ormina and velasquezi groups. These are most convincingly defined by differences in the female genitalia. Of these, the ormina group is the more variable and the placement of M. rodmani Blahnik and Holzenthal and M. simplicis , n. sp. within this group is conjectural, since females are not known for these species. Some species in both the ormina and velasquezi groups have modified scale-like setae paralleling the major veins of the fore- and/or hind wings. A similar development occurs in a few species of the leroda group of the subgenus Mortoniella . The character is mostly hidden by the normal setae present and may not be readily evident. The scales seem to be readily lost in alcohol preserved specimens. In some species of the velasquezi group, there is a more general and readily apparent field of scale-like setae on the hind wing. These are of somewhat different morphology than the scale-like setae paralleling the veins (stalked basally and upright, rather than leaf-like and flattened). Whether the presence of scale-like setae represents an independent character development in the two groups, or an apomorphy for the group, lost in some taxa, is uncertain.
Adult —Length of forewing: 1.7-4.0 mm; females slightly larger than males. Forewing with forks I, II, and III, forks I and II usually sessile, sometimes with fork II stalked ( Fig. 104A View Figures 102-104 ); hind wing with fork II only ( Fig. 102B, 103B, 104B View Figures 102-104 ), basal forks of Rs and M veins at about midlength and narrowly separated (ormina group), or with basal fork of Rs more proximal (velasquezi group). Crossveins of forewing (r, s, r-m, m, m-cu, cu) linear or nearly so and usually hyaline; hind wing with r-m only (ormina group, Fig. 103B, 104B View Figures 102-104 ), or with crossveins absent (velasquezi group, Fig. 102B View Figures 102-104 ). Costal margin of hind wing without distinct inflection, wings more or less spear-shaped. Forewing rather broadly rounded apically in velasquezi group ( Fig. 102A View Figures 102-104 ), narrow and angulate in ormina group ( Fig. 103A, 104A View Figures 102-104 ), apex of hind wing angulate. Fore- and/or hind wing frequently with adpressed scale-like setae paralleling veins; some species of velasquezi group also with patch of upright scale-like or hair-like setae on hind wing. Spur formula 0:3:4 (0:2:4, as variant in M. usseglioi ). Overall color varying from light brown to dark brown; forewing usually marked with white or whitish setae at anastamosis, in some species of ormina group with small white spots apically.
Male genitalia —Ventral process of segment VI relatively elongate, narrow, and posteriorly projecting (length usually 2 times width or more). Segment IX with anterolateral margin rounded and produced in ventral ½, posterolateral margin nearly straight; segment deeply mesally excised dorsally and ventrally, forming lateral lobes, lobes usually separated dorsally by ½ or more of width of segment. Tergum X well developed, setose, usually with deep V-shaped or U-shaped apicomesal invagination ( M. aequalis an exception); apicolateral lobes usually simple in structure and not strongly sclerotized. Inferior appendages directly fused to one another and to ventral margin of phallic ensemble, generally short and reduced in ormina group, or in velasquezi group with more distinct dorsolateral process and apex sometimes elongate and fused to mesal spinelike process; appendages always with paired mesal pockets accommodating rod-like projections of phallobase, these very enlarged in velasquezi group, apices of pockets with relatively short to very elongate spine-like projections, typically narrow in ormina group, widened in velasquezi group. Basal segments of parameres variable, usually relatively simple in structure in velasquezi group (short, rounded and fused to one another), often more modified in ormina group, sometimes with elongate rod-like projections; apical segments (paramere appendages) usually present, short to elongate, typically more or less rod-like, often strongly curved. Phallobase short or very short, always with, paired rod-like projections from the apicoventral margin, these short in ormina group (sometimes with apex flared), long and with apices flared in velasquezi group; dorsal margin with moderately elongate mesal spine, relatively simple in structure in some, more often with lateral margins projecting; dorsal phallic spine without basal articulation with phallicata, apex strongly reflexed in most. Phallicata usually short and only indistinctly sclerotized, with distinctly sclerotized, rounded, lateral projection in velasquezi group. Endophallic membrane very variable, usually relatively short and simple in structure, sometimes with dorsal or lateral spines or spine-like structures or with membranous lateral lobes; in velasquezi group with pair of ventral sclerites or spine-like projections and dorsomesal projection, typically with small spines or short spine-like projection; phallotremal spines not usually evident ( M. collegarum a notable exception).
Etymology —This subgenus is named Nanotrichia from the Greek words nanos, meaning small, and referring to the small size of the species in the subgenus, and trich - for hair, a term generally applied to species in the order Trichoptera because of their hairy wings. The name is feminine in gender.
| A |
Harvard University - Arnold Arboretum |
| V |
Royal British Columbia Museum - Herbarium |
| I |
"Alexandru Ioan Cuza" University |
| M |
Botanische Staatssammlung München |
| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.