
Atanatolica bonita, Costa, Anne M. & Calor, Adolfo R., 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3790.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:B808291C-8450-43A8-904D-192C24C494C5 |
|
DOI |
https://doi.org/10.5281/zenodo.6123010 |
|
persistent identifier |
https://treatment.plazi.org/id/03F78798-FFA7-FF9A-30E5-FBA4FAF41A1C |
|
treatment provided by |
Plazi |
|
scientific name |
Atanatolica bonita |
| status |
sp. nov. |
Atanatolica bonita new species
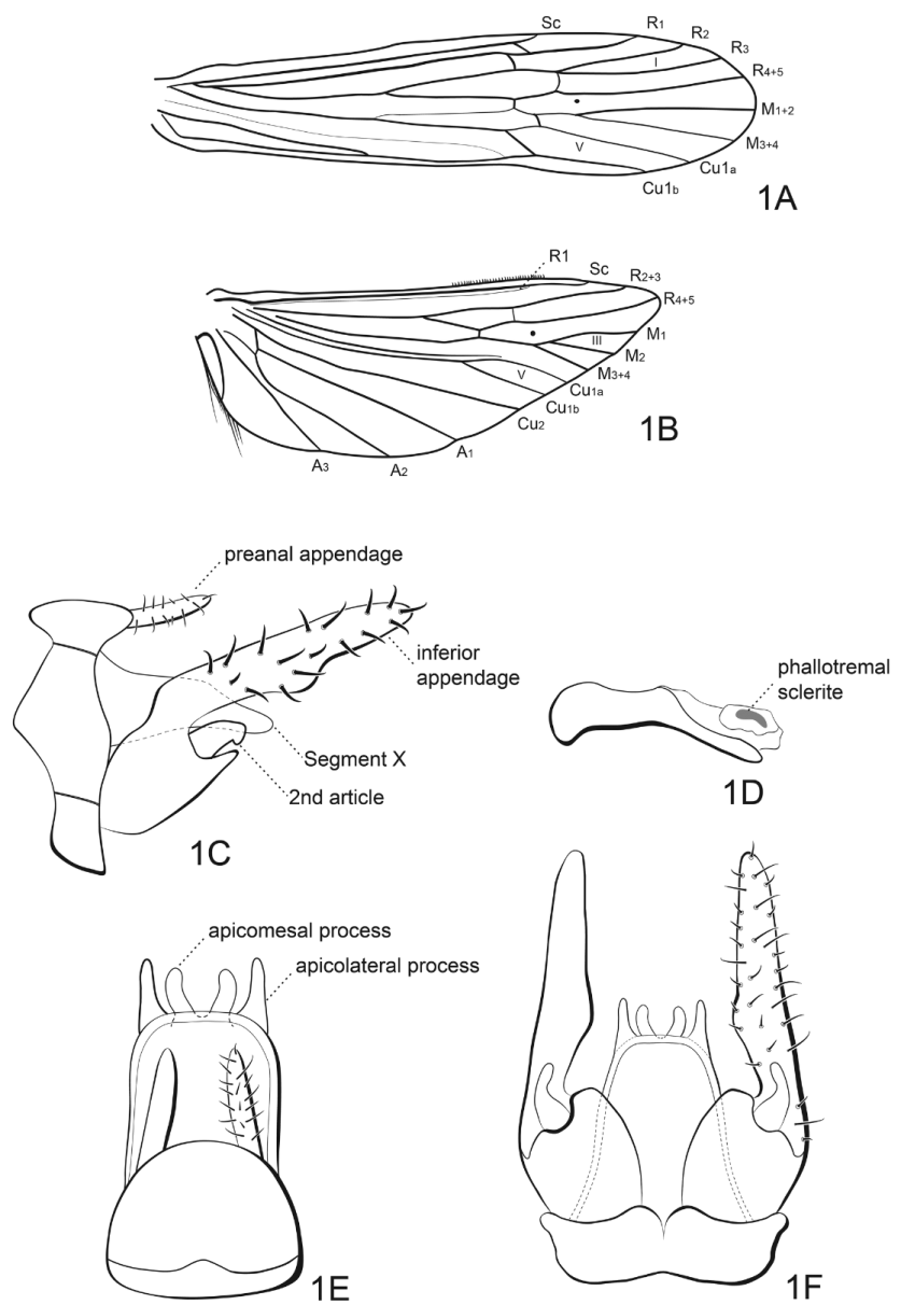
( Figs. 1–2 View FIGURE 1 View FIGURE 2 )
Diagnostic characters. The male can be easily distinguished from its congeners by possessing two apicomesal and two long, digitate, subequal apicolateral processes on segment X and a well-developed, long, thumb-shaped second article of each inferior appendage. The female can be diagnosed by the presence of a funnel-shaped vaginal apparatus.
Adult: Forewing length 7–9 mm; hind wing length 5–7 mm (n = 10).
Head brown (in alcohol) with white and brown setae (pinned). Frontal setal wart oval; malar space on each side narrow. Antennae long, with white setae at the base of each flagellomere; brown, about twice body length. Maxillary palps brownish, covered by brown setae, 5-segmented.
Thorax brown with many white setae. Wings brownish with brown and golden setae. Vein pattern typical of genus: Forewing forks I and V present in male ( Fig. 1 View FIGURE 1 A); forewing forks I, III and V present in female. Hind wing forks III and V present in both sexes ( Fig. 1 View FIGURE 1 B). Tibial spur formula 0,2,2.
Male: Mean forewing length 9.1 mm ( 8.5–9.2 mm); mean hind wing length 6.6 mm ( 6.4–6.8 mm) ( Fig. 1 View FIGURE 1 ). Mean body length 6.5 (6.1–7.0 mm) (n = 15).
Genitalia having abdominal segment IX, in lateral view, narrow, sutures separating each side of segment in 3 parts and posterior margin projected medialy. Preanal appendages shorter than segment X, with several setae ( Figs. 1 View FIGURE 1 C, 1E). Tergum X with 1 pair of apicomesal processes and 1 pair of apicolateral processes long and digitate with subequal lengths ( Figs. 1 View FIGURE 1 E, 1F). Inferior appendages densely covered by setae; basal portion of each inferior appendage with apicomesal process best seen in lateral view ( Fig. 1 View FIGURE 1 C); second article of each inferior appendage prominent, broad, thumb-shaped ( Figs. 1 View FIGURE 1 C, 1F). Phallic apparatus narrow gradually curved ventrad in apical half, with sclerite ( Fig. 1 View FIGURE 1 D).
Female: Mean forewing length 6.9 mm ( 6.9–7.9 mm); mean hind wing length 4.8 mm ( 4.8–6.5 mm). Mean body length 5.2 ( 5.2–5.6 mm) (n = 6).
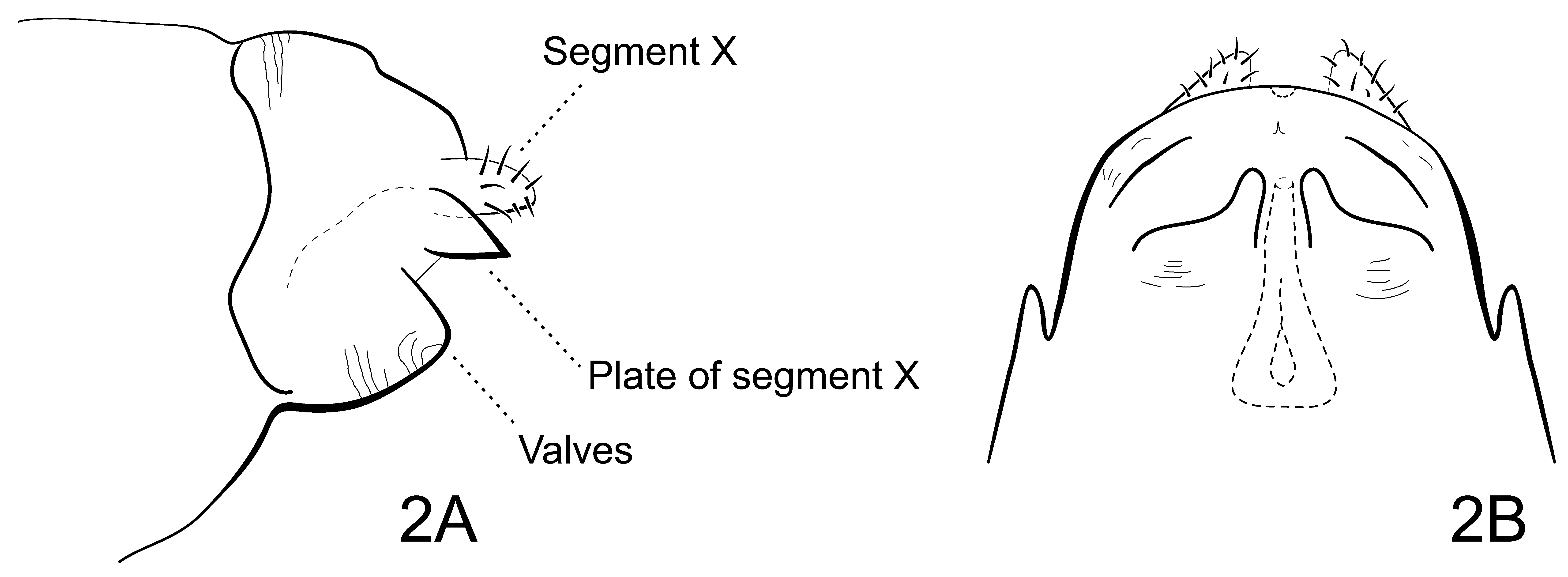
Genitalia having abdominal segment IX broad, U-shaped and grooved. Appendages of segment X short, slender, with setae; segment X bearing thin, transverse sclerotized plate, projecting below appendages X and dorsal of valves and sternum IX. Valves small, with pair spine-like processes above them ( Fig. 2 View FIGURE 2 A). Vaginal apparatus funnel-shaped ( Fig. 2 View FIGURE 2 B).
Egg, Larva and Pupa: Unknown.
Material examined. Holotype ♂ (alcohol): Brazil: Bahia, Camacan, RPPN Serra Bonita , stream 3, 15°23’02”S, 39°34’10”W, 806 m, 01.viii.2008, A. Calor, L. Lecci, L.C. Pinho & R. Moretto leg. ( MZUSP).
Paratypes: same as holotype, 0 2 ♂ (pinned) and 0 1 ♀ ( MZUSP); same, except aerial entomological net, 0 3 ♂ (pinned); same, except 14.iv.2008, 0 3 ♂; same, except 02.viii.2008, 0 2 ♂ and 0 1 ♀; same, except 15°23’10”S, 39°34’03”W, 819 m, aerial entomological net, 0 2 ♀; same, except 04.viii.2008, 0 3 ♂ ( UFBA); same, except 25.ix.2008, 0 1 ♂ and 0 1 ♀; same, except 19°22’57”S, 39°33’21”W, 04.viii.2008, 0 1 ♂ (pinned) ( UMSP).
Supplementary material: same as holotype, 20 ♂ and 0 5 ♀; same, except 01.x.2008, 13 ♂ and 01♀; same, except 01.xi.2008, 10 ♂; same, except 01.xii.2008, 0 8 ♂; same, except 01.i.2009, 0 2 ♂ and 0 2 ♀; same, except 01.ii.2009, 11 ♂ and 0 1 ♀; same, except 01.iii.2009, 0 2 ♂ and 0 1 ♀; same, except 01.iv.2009, 0 2 ♂; same, except 01.vi.2009, 0 2 ♂ and 0 2 ♀; same, except 01.vii.2009, 0 5 ♂ and 0 5 ♀; same, except 01.viii.2009, 16 ♂ and 0 4 ♀; same, except 01.ix.2009, 22 ♂ and 0 2 ♀; same, except 01.x.2009, 15 ♂ and 0 6 ♀; same, except 01.xi.2009, 20 ♂ and 0 3 ♀; same, except 01.xii.2009, 24 ♂ and 0 2 ♀; same, except 01.iii.2010, 0 3 ♂; same, except 01.i.2011, 0 1 ♂; same, except 01.iii.2011, 12 ♂ and 0 4 ♀; same, except 01.iv.2011, 24 ♂, 0 6 ♀; same, except 01.vi.2011, 11 ♂ and 0 2 ♀; same, except 01.vii.2011, 0 5 ♂, 0 2 ♀; same, except 01.viii.2011, 26 ♂, 0 6 ♀; same, except 01.x.2011, 0 8 ♂, 0 4 ♀; same, except 15º23’1.4’’S, 39º34’10’’W, 25.xi.2011, U.V./White light, R. Mariano, F. Quinteiro, T. Duarte, & E. Silva, E. legs., 0 2 ♂.
Distribution. Brazil (BA).
Etymology. The specific epithet, bonita , refers to the type locality of the species, RPPN Serra Bonita . Additionally, the word “ bonita ” is the Portuguese adjectival equivalent to good-looking, which is indeed an attribute of both the reserve and the species.
Taxonomical remarks. According to Holzenthal (1988) the genus can be subdivided in two groups: The A. dominicana Group with forewing fork I petiolate, and the A. brasiliana Group with forewing fork I sessile. This latter character is shared among all members of the A. brasiliana Group, including A. bonita n. sp.. Concerning the hind wings, A. bonita n. sp. is very similar to A. brasiliana , but differs from A. flinti ( A. brasiliana Group). Atanatolica flinti has an open discoidal cell ( rs crossvein absent) and fork I is present ( Holzenthal 1988).
In general, A. bonita n. sp. is more similar to A. brasiliana than to A. flinti , considering the wings patterns and morphology of genitalia. The new species and A. brasiliana share the following characters: Preanal appendages shorter than segment X, segment X with apicomesal processes, and similar shapes of the second article of each inferior appendage. However, A. bonita n. sp. can be distinguished from A. brasiliana based on the long apicolateral processes X, the long second article of each inferior appendage, and the broad phallic apparatus gradually curved ventrad in its apical half (versus short apicolateral processes X, short second article of each inferior appendage, and more slender phallic apparatus in A. brasiliana ).
Biological remarks. Malaise traps were positioned near streams in four localities: 15°23’28”S, 39°33’56”W, 830 m (stream 1, tributary of Panelão River); 15°23’26”S, 39°33’58”W, 828 m (stream 1, tributary of Panelão River); 15°23’37.3”S, 39°33’51.4”W, 814 m (stream 1, tributary of Panelão River), and 15°23’03”S, 39°34’00.1”W, 806 m (stream 3, first order tributary of Braço do Sul River). The majority of the specimens of A. bonita n. sp. were collected from stream 3 between August 2008 and December 2011. Only seven specimens were collected in light traps, probably because of their diurnal activity ( Flint et al. 1999).
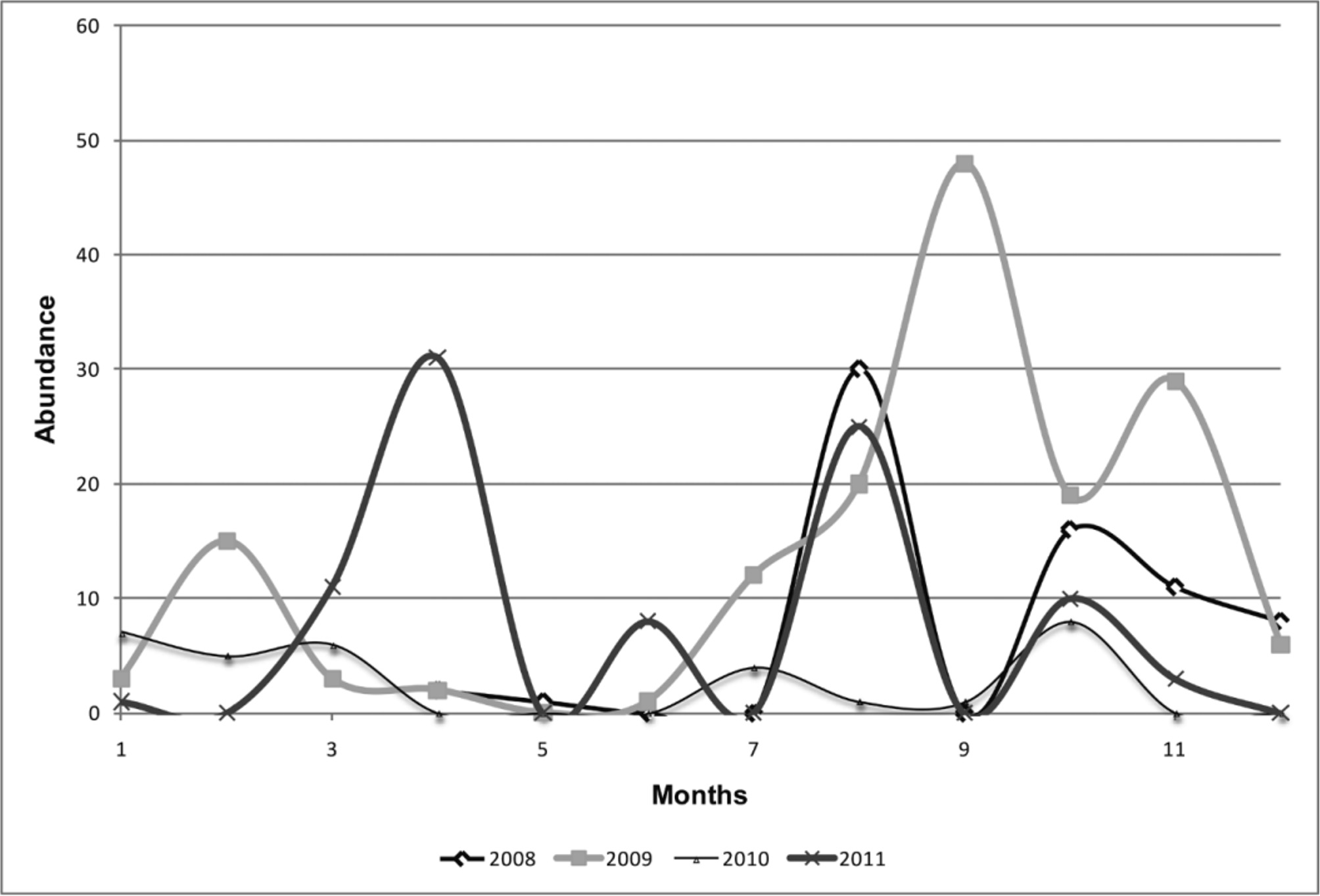
Considering the specimens collected from Malaise traps (n = 343), we observed a trend for adults to occur in specific periods of the year. A scarcity of published phenological data, especially for Neotropical caddisflies, makes these data for the new species especially interesting.
During these years, 2009 was the year in which the largest number of specimens was collected, reaching almost 50% of all the material sampled. In 2009 the species is more abundant in the second semester, numbers collected increased in August with a gradual decrease from October onwards ( Fig. 3 View FIGURE 3 ). This pattern was not observed in 2011, with a high number of specimens collected in April, also.
In the Neotropical Region, Wolf et al. (1988) proposed a negative relationship between emergence of aquatic insects and periods with high precipitation; temperature and rainfall seem to be the main regulatory factors. In this case, the high emergence of the new species (as indicated by the number of collected specimens) in the second semester, especially from July to August can be explained by lower temperature and precipitation. On the other hand, the decrease in abundance can be understood by higher temperature and precipitation with the onset of the wetter season. According to Liuth et al. (2013), these two seasons can be defined in the study area: A wetter season (from November to April) with high temperatures and elevated precipitation, and a drier season (from May to October) with low temperatures and light precipitation.
Flint and Masteller (1993) observed that increase in water volume interferes with Trichoptera emergence, as the immatures may be killed. In this context, the reduction in abundance during the rainy season is a well documented phenomenon ( Bispo & Oliveira 1998; Diniz-Filho et al. 1998; Bispo et al. 2001; 2004; 2006), especially in first order streams. Rainfall interacts with local factors such as topography and riparian characteristics (e.g., with or without vegetative cover); and variations of current speed, water volume, and dissolved oxygen, among other factors, to cause dramatic impacts on the aquatic insect fauna ( Yokoyama et al. 2012). Thus, seasonal variation in the precipitation regime could possibly be responsible for demographic differences during the year.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |