
Tyrannosaurus rex (Osborn, 1905)
|
publication ID |
https://doi.org/ 10.2110/palo.2009.p09-056r |
|
persistent identifier |
https://treatment.plazi.org/id/03F987D8-5A3E-FFFB-D9E6-FC051AF4FC10 |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
Bite marks preserved on bone are not uncommon in the vertebrate fossil record and have the potential to yield insights into aspects of behavioral evolution and ecology ( Buffetaut, 1983; Currie and Jacobsen, 1995; Erickson et al., 1996; Tanke and Currie, 1998; Hanna, 2002; Avilla et al., 2004; Katsura, 2004). Determining whether bite marks and other injuries preserved on fossil bones are the result of interspecific or intraspecific conflict, predation, post-mortem scavenging or disease, however, is often difficult. Furthermore, positive identification of the species that caused the injury and inferences about behavior from such traces are far more challenging tasks. Here we report a juvenile tyrannosaurid skeleton ( BMR P2002.4.1 ) that exhibits a series of partially healed lesions along the left maxilla and nasal, which we interpret as a bite mark injury. By utilizing the Extant Phylogenetic Bracket (EPB) method ( Witmer, 1995), we interpret this injury as resulting from intraspecific aggression between two similarsized juvenile tyrannosaurids and discuss the insights the specimen provides for juvenile tyrannosaurid behavior.
GEOLOGIC SETTING
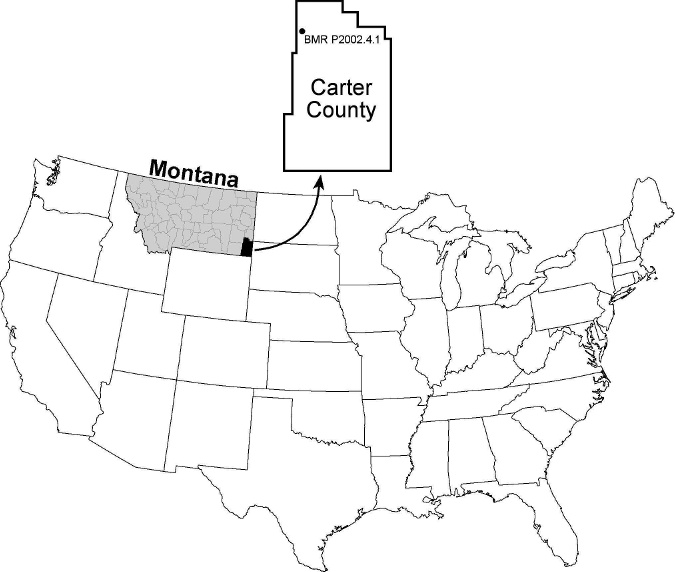
Specimen BMR P2002.4.1 was collected from the upper half of the Hell Creek Formation of western Carter County, Montana in 2002 ( Fig. 1 View FIGURE 1 ) and is latest Maastrichtian in age. Exact coordinates of the collection site are on file in the paleontology collections at the Burpee Museum of Natural History, Rockford, Illinois (BMR), where the specimen is housed. The tyrannosaurid skeleton was buried in a lenticular clay-ball conglomerate unit composed of poorly sorted sand, silt, rounded green clay rip-up clasts, and diagenetically produced siderite nodules that partly encased several bones. The bone-bearing unit displayed a rapid lateral variation in thickness from 12 to 40 cm and was overlain by a massive, poorly cemented, cross-bedded sandstone ( Henderson and Harrison, 2008).
In southeastern Montana the Hell Creek Formation averages 150 meters in thickness ( Johnson et al., 2002) and contains a diverse and well-preserved latest Cretaceous biota. The formation is especially well known for its fossil vertebrates, which include fish, amphibians, aquatic and terrestrial reptiles, mammals, dinosaurs, and rarely pterosaurs ( Fastovsky, 1987; White et al., 1998; Johnson et al., 2002; Henderson and Peterson, 2006). A diverse and well-preserved fauna and flora was recovered from the BMR P2002.4.1 excavation site ( Harrison et al., 2006).
DESCRIPTION
BMR P2002.4.1 consists of 145 bones of a small tyrannosaurid, approximately seven meters in length. Based on histological characters, degree of skeletal fusion, and thin-walled bone structure, BMR P2002.4.1 is considered a late juvenile animal rather than a subadult since it had not yet reached a phase of rapid growth ( Erickson et al., 2006; Henderson and Harrison, 2008).
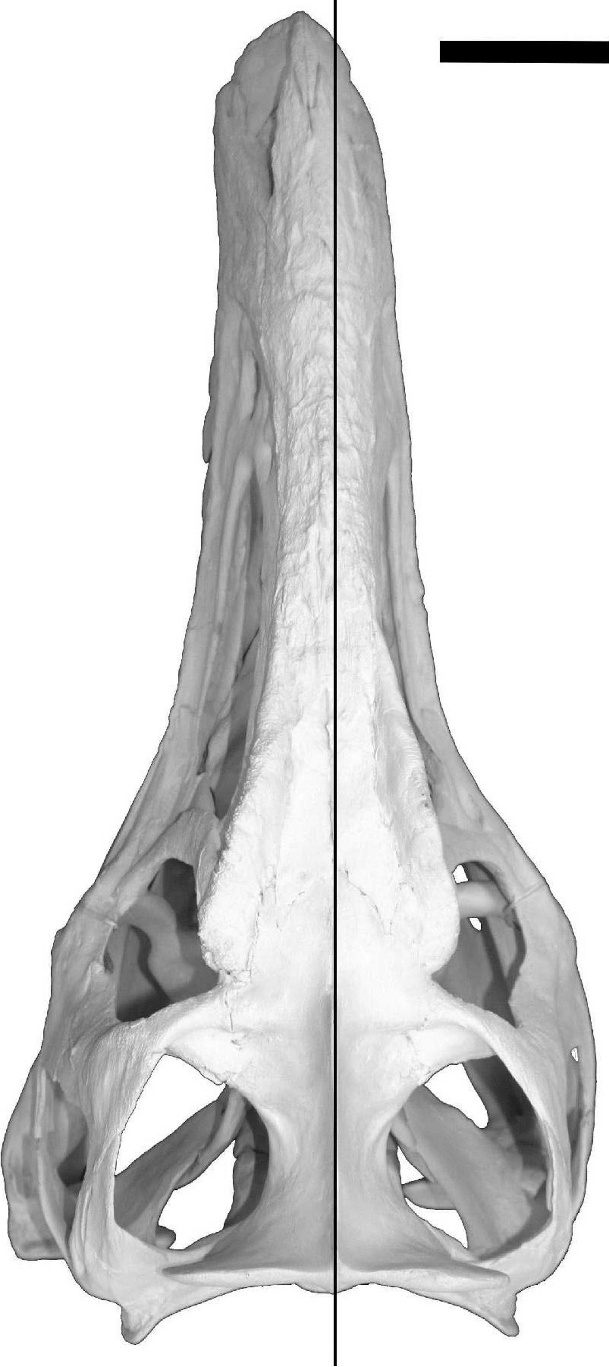
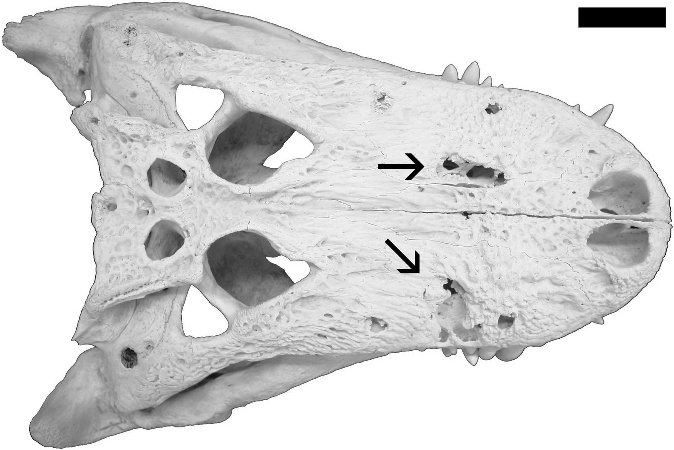
The skeleton was partially articulated when discovered, and consists of a nearly complete and well-preserved skull, found disarticulated above the animal’s hips, and a large portion of the postcranial skeleton. The depositional setting indicates rapid burial of the skeleton, and there is no evidence for predation or extensive scavenging. Cranial characters closely resemble those of another late Maastrichtian specimen (CMNH 7541 [Cleveland Museum of Natural History]), which has been interpreted as belonging to either a juvenile Tyrannosaurus rex ( Carr, 1999; Carr and Williamson, 2004) or a separate taxon, Nanotyrannus lancensis ( Bakker et al, 1988; Currie, 2003a; 2003b). Research is currently ongoing into the systematic position of both specimens. Four lesions are present on the left side of the skull of BMR P2002.4.1 ( Fig. 2 View FIGURE 2 A–B). The lesions are lenticular shaped, rostrocaudally long, and fully penetrate the left nasal and maxilla, which is 6 mm thick near the puncture marks. Several of the preserved lesions display bone repair (cicatrisation), indicating that the animal survived these injuries to its skull. There is no osteological evidence for infections at these puncture sites.
The anteriormost puncture is located ~15 mm posterior to the left bony naris, at the nasomaxillary suture ( Fig. 2 View FIGURE 2 C–D). This puncture has a maximum length of 15 mm and a maximum height of 5 mm. The edges are fractured with partial bone repair. The left maxillary process of the nasal is absent, apparently excised during the injury ( Fig. 2C View FIGURE 2 ). Three additional lesions are preserved on the left maxilla ( Fig. 2 View FIGURE 2 E–F). The rostralmost puncture is present directly above the maxillary fenestra and is 10 mm in length and 9 mm in maximum height. The edges are fractured without evidence of cicatrisation. The second puncture is positioned 8 mm posterior to the first and is 18 mm in length and 4 mm at maximum height. The edges of the puncture consist of smooth cortical bone. The third puncture is 12 mm posterior to the second hole and is 19 mm in length and 5 mm at maximum height; its edges are also of smooth cortical bone.
In addition to the lesions, the reconstructed skull of BMR P2002.4.1 demonstrates abnormal asymmetry, with the distal portion of the muzzle curved laterally to the left side ( Fig. 3 View FIGURE 3 ). The curvature is not considered severe enough to have affected the animal’s ability to bite or capture prey. Post-depositional deformation is a common cause of bone distortion in fossil specimens, but the asymmetry of this specimen is considered primary—that is, caused by the injury, with the distortion propagated by subsequent growth—rather than a taphonomic feature. When discovered, the skull of BMR P2002.4.1 was disarticulated with its nasal lying dorsoventrally and both maxillae lying on their lateral sides. The nasal and maxillae fit well along their articular surfaces despite the disarticulation, and there is little evidence of postdepositional deformation in any of the other preserved cranial elements. This strongly suggests that the lateral curvature of the muzzle is the result of facial injury rather than diagenetic deformation. Furthermore, there is no sedimentological evidence of post-depositional compaction; the clay balls in the conglomerate that encase the bones are spherical, not compressed to oriented lenticular clasts.
A major pathology with evidence of active infection at the time of death is present on the first phalanx on the second pedal digit of the left pes (C.P. Vittore, personal communication, 2009). There is no evidence to suggest, however, that this pathology is related to the facial injuries.
DISCUSSION
There is little doubt that the loss of the left lateral process of the nasal and the lesions to the left maxilla of BMR P2002.4.1 occurred at the same time. This is indicated by the uniform shape and size of the lesions and the presence of healed bone tissue along their edges. In addition, the distribution of the lesions along a gently arcuate line and the similar degree of healing supports the hypothesis that these injuries were inflicted simultaneously, probably from a bite to the face. The lateral curvature of the muzzle described above likely resulted from differential growth of the face following the loss of the left lateral nasal process.
The only fossil vertebrates preserved in the Hell Creek Formation that are large enough to puncture the skull of BMR P2002.4.1 are crocodilians and theropod dinosaurs. The lenticular-shaped lesions are inconsistent with the circular cross sections of crocodilian teeth, but they are consistent with the lenticular cross sections of juvenile theropod teeth. The maxillary teeth of BMR P2002.4.1 are laterally compressed; a characteristic of juvenile tyrannosaurids in general. The size, shape, and spacing of the maxillary teeth of BMR P2002.4.1 correspond well with the size, shape and spacing of the lesions, indicating that this animal was bitten by another juvenile tyrannosaurid of its size. The location and spacing of the facial injuries indicate that the bite was inflicted by the grazing anterior maxillary teeth of an adversary, which was in front of and facing BMR P2002.4.1 ( Fig. 4 View FIGURE 4 A– B). The smooth cortical bone covering the edges of the lesions indicates that the bite was not fatal and healing was well underway or complete at the time of death.
The lesions in BMR P2002.4.1 provide insight into the social behavior of juvenile tyrannosaurids. Evidence for face biting is not uncommon in theropod dinosaurs ( Tanke and Currie, 1998). While intraspecific predation and cannibalism have been previously suggested in tyrannosaurids ( Tracey, 1976; Naeye, 1996), true predation and cannibalism usually involve a much larger individual attacking a smaller one ( Tanke and Currie, 1998) and are not likely scenarios for the bite described here.
While social behavior cannot be directly observed in or unequivocally inferred from the fossil record, it can be inferred from evidence of eating and from injuries ( Tanke and Currie, 1998; Avilla et al., 2004; Katsura, 2004). The ability of tyrannosaurids to puncture and crush bone is well documented ( Abler, 1992; Erickson and Olson, 1996; Erickson et al., 1996; Hurum and Currie, 2000; Meers, 2002), and evidence of feeding is relatively common at fossil vertebrate sites ( Chure et al., 1998). Behavior can also be inferred using the Extant Phylogenetic Bracket (EPB) method ( Witmer, 1995). This method is generally used for soft-tissue reconstruction and behavioral inference by comparing the osteological correlates of living members of a group to infer a characteristic in an extinct member. The EPB has previously been used to interpret nesting and parental behavior in non-avian theropods ( Varricchio et al., 2008), using extant crocodilians and birds as living members of Archosauria. A similar model was used for the purposes of this study, where intraspecific aggressive behaviors were investigated in extant crocodilians and birds.
Puncture marks along the maxilla and nasal, and skewed jaws similar to the injuries on BMR P2002.4.1 are commonly seen on extant crocodilians as the result of non-fatal intraspecific aggression ( Kalin, 1936), especially on the saltwater crocodile, Crocodylus porosus ( Webb and Messel, 1977). Extant crocodilians display a variety of complex social behaviors that often result in intraspecific aggression due to conflicts in dominance hierarchies, territoriality, or courtship ( Lang, 1989). The injury described here lacks evidence of the shaking or rolling behaviors that extant crocodilians commonly display with prey capture. The lesions present on BMR P2002.4.1 are consistent with injuries in extant crocodilians exhibiting intraspecific aggression (i.e., head-slapping) rather than prey capture (rolling, shaking). Because the puncture wounds present on BMR P2002.4.1 are similar to those found on extant crocodilians ( Fig. 5 View FIGURE 5 ), we consider it likely that they both resulted from roughly comparable intraspecific aggressive behavior.
Erickson at al. ( 2006) indicate that Tyrannosaurus rex reached sexual maturity at 14 years of age. BMR P2002.4.1 has been histologically determined to be 12 years old at the time of death ( Erickson et al., 2006). Because BMR P2002.4.1 was probably not sexually mature, we suggest that the injury did not likely result from courtship behavior.
Aggression in dominance hierarchies and territoriality between extant crocodilians commonly result in conflict and injury ( Webb and Messel, 1977). Territoriality is most common in small groups, however, whereas dominance hierarchies are typically seen in large communities and involve more massive individuals ( Lang, 1989). Saltwater crocodiles are territorial and aggressively display and vocalize during confrontations, especially between two males, and these conflicts can include biting the face of an opponent. This aggression may be brought on by the defense of foraging areas, basking locations, access to mates, or a combination of these factors ( Lang, 1989). We speculate that the facial injury of BMR P2002.4.1 was the result of a territorial or dominance bout, similar to those observed in extant crocodilians.
Blanco et al. (1997) documented inter- and intraspecific aggression in cinereous vultures ( Aegypius monachus ) and griffon vultures ( Gyps fluvus ). Aggressive behaviors are commonly intraspecific in griffon vultures, though conflicts rarely result in serious injury. The most violent interactions, where injuries are more likely, however, usually involve juveniles ( Blanco et al., 1997).
While juvenile tyrannosaurids are both rare and poorly known with little evidence upon which to infer their behavior, injuries that have been phylogenetically determined as bite marks suggest behaviors such as intraspecific aggression. Placement of BMR P2002.4.1 on a logistic growth curve for Tyrannosaurus rex indicates that the specimen was not yet sexually mature ( Erickson et al., 2006), suggesting that this behavior was not related to courtship or mating and that juvenile tyrannosaurids engaged in agonistic behavior prior to reaching sexual maturity.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |