
Coleophora ericarnella Baldizzone
|
publication ID |
https://doi.org/10.11646/zootaxa.4111.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:9F38106A-9396-4CE9-990D-553EEE37C59A |
|
DOI |
https://doi.org/10.5281/zenodo.5689373 |
|
persistent identifier |
https://treatment.plazi.org/id/95EC2CDC-BAFD-4E5E-B8DA-3E6784BB26F2 |
|
taxon LSID |
lsid:zoobank.org:act:95EC2CDC-BAFD-4E5E-B8DA-3E6784BB26F2 |
|
treatment provided by |
Plazi |
|
scientific name |
Coleophora ericarnella Baldizzone |
| status |
sp. nov. |
Coleophora ericarnella Baldizzone , sp. nov.
Type material. HOLOTYPE ♂ “Italien—Friuli | Alta Val Torre | Vedronza | 350 m | 24.6.2004 —LF | leg. H. Deutsch” “Bldz PG n° 14824”, coll. MFSN.
PARATYPES: 8 ♂♂ (PG Bldz 14874, 14875, 15708, 15709), idem; 1 ♂, idem (CNCLEP00068771), coll. CNC; 2 ♂♂ (PG Bldz 14484, 14485) “Italien—Friuli | Valle di Musi | C.re [= Casera] Tanatcason | 750 m | 26.6.2004 —LF | leg. H. Deutsch”, in coll. MFSN, coll. Deutsch, coll. Baldizzone; 1 ♀ (PG Wolf 11947) “ Italien / Friaul, Val Cellino/Val Verron 600 m. / 20.06.2013 / leg. J. Viehmann”, coll. van der Wolf; 1 ♂ “ Italy, South Tyrol, Cimabanche/Schluderbach, 1600 m, 21.7.2013, leg. Huemer”, ( TLMF Lep 12416) coll. TLMF; 1♂ (PG Bldz 14785) “ Italia; Trentino; TN, Fiera di Prim.[iero], Val Canali, Villa Welsperg | 1050 m | 19.VI.2009 | leg. Timossi G.”, coll. Timossi; 2 ♂♂ (PG Bldz 1045, 14503) “Trentino | Val Sarca, Pietramurata | con luce | 250 m | 13.VII. [19]58 | E. Jäckh”, 1 ♂ (PG Bldz 922), ibidem, 4.VII.1959, 1 ♂ (PG Bldz 1489), ibidem, 10.VII.1960, coll. Baldizzone; 1 ♂, ibidem, 16.5.1961, K. Burmann leg., coll. TLMF; 1 ♂ (PG Bldz 11244) “ Italia | Trentino | Südlich Nago | 22.6.1987, leg. R. Keller”, coll. Baldizzone; 1 ♀ (PG Bldz 5613) “ Italia sept. | Trentino | Villamontagna | 600 m | 18.6.1982 | K. Burmann leg.”, coll. Baldizzone. 7 ♂♂ (PG Bldz 14491, 14877) “Austria—Osttirol | Nörsach, Rabantberg | Felsenhang | 630 m | 07.6.2003—LF | leg. H. Deutsch”; 1 ♂ Austria—Osttirol, Nörsach, Rabantberg, Felsenhang, 630 m, 3.6.2002, leg. H. Deutsch ( TLMF Lep 17598), coll. TLMF; 1 ♂ “Austria—Osttirol | Lavant, Kienbichl | 750 m | 10.6.2003 —LF | leg. H. Deutsch”; 1 ♂ (PG Bldz 15374) “Austria—Osttirol | Lavant, Kienbichl | 950 m | Waldsteppen, Felsen | 13.7.2009 | leg. H. Deutsch”; 2 ♂♂ Austria—Osttirol, Lavant, Lavanter Almtal, 1200 m, 20.6.2002, leg. H. Deutsch ( TLMF Lep 17596, TLMF Lep 17597), coll. TLMF; 1 ♂ “Austria—Osttirol | Lavant, Forchach | 650 m | 17.6.2003 —LF | leg. H. Deutsch”; 4 ♂♂ (PG Bldz 14876, 15710) “Austria—Osttirol | Lavant | Lavanter Forchach | 650 m | 22.6.2005 | leg. H. Deutsch”; 1 ♂, ibidem, 24.6.2008, leg. H. Deutsch (CNCLEP00068770), coll. CNC; 13 ♂♂, ibidem, 24.6.2009, leg. H. Deutsch; 2 ♂♂ (PG Bldz 14828), 4 ♀♀ (PG Bldz 14822, 14827), ibidem, 14.6.- 10.7.2009, ex larva Erica carnea , leg. H. Deutsch; 1 ♂, 4 ♀♀ (PG Bldz 15372, 15373), ibidem, 27.5.2011, ex larva Erica carnea , leg. H. Deutsch, coll. Deutsch et coll. Baldizzone; 1 ♂ Italy, Suedtirol, N Völser Weiher/ Völs am Schlern, 1040 m, 15.6.2006, leg. P. Huemer ( TLMF Lep 18592), coll. TLMF; 1 ♂ (PG Wolf 6640) “ Jugoslavija, Slovenien, Triglav Vrata, 1100 m, 27.VII.1984, K. Schnack”, coll. ZMUC.
28 ♂♂, Italia, Trentino Val Sarca, Pietramurata, at light, leg. E. Jäckh, various dates as follows: 1 ♂, 5.VII.1958 (USNMENT01200067); 2 ♂♂, 19.VII.1958 (USNMENT01200063, USNMENT01200064); 2 ♂♂, 2.VII.1959 (USNMENT01200075, USNMENT01200079); 3 ♂♂, 4.VII.1959 (USNMENT01200074, USNMENT01200077, USNMENT01200078); 1 ♂, 10.VII.1959 (USNMENT01200076); 1 ♂, 25.VII.1959 (USNMENT01200068); 1 ♂, 10.VII.1960 (USNMENT01200065); 1 ♂, 14.VII.1960 (USNMENT01200069); 2 ♂♂, 17.VII.1960 (USNMENT01200060, USNMENT01200061); 2 ♂♂, 22.VII.1960 (USNMENT01200058, USNMENT01200059); 1 ♂, 26.VII.1960 (USNMENT01200066); 1 ♂, 10.VII.1961 (USNMENT01200085); 2 ♂♂, 7.VII.1970 (USNMENT01200070, USNMENT01200071); 1 ♂, 8.VII.1970 (USNMENT01200072); 4 ♂♂, 29.V.1977 (USNMENT01200073, USNMENT01200082, USNMENT01200083, USNMENT01200084); 2 ♂♂, 29.VII.1978 (USNMENT01200080, USNMENT01200081); coll. USNM.
Additional specimen examined. 1 ♂ (PG Bldz 3489) “Trentino, Val Sarca, Pietramurata, 200 m, al lago con luce, 15.VII.1958, E. Jäckh”, “sarcaella Toll”, coll. ISEZ.
Diagnosis. Coleophora ericarnella is smaller than C. pyrrhulipennella ( Figs. 3–4 View FIGURES 1 – 7 ) and C. pulchripennella ( Figs. 5–6 View FIGURES 1 – 7 ), especially the females. The coloration is overall paler, ochre instead of brown, more golden yellow, with the white streaks thinner and less prominent. In the male genitalia, the cucullus is shorter and thicker, the ventral edge of sacculus is more straight and almost without a protuberance at the ventral angle, which is distinct in C. pyrrhulipennella ( Fig.15 View FIGURES 13 – 16 ) and even more so in C. pulchripennella ( Fig. 16 View FIGURES 13 – 16 ); the cornuti are smaller and less numerous. In the female genitalia, the most distinctive feature is the spinulose section of the ductus bursae which is shorter than in C. pyrrhulipennella ( Fig. 18 View FIGURES 17 – 22 ), and very thin as opposed to being very thick in C. pulchripennella ( Fig. 19 View FIGURES 17 – 22 ). The signum of C. ericarnella ( Fig. 20 View FIGURES 17 – 22 ) has a more robust spine with a smaller, less expanded and unnotched base, whereas that of C. pyrrhulipennella ( Fig. 21 View FIGURES 17 – 22 ) has a wide base with a medial notch; in C. pulchripennella ( Fig. 22 View FIGURES 17 – 22 ), the signum base is evenly rounded and wider than in C. ericarnella .
Description. ♂ ( Fig. 1 View FIGURES 1 – 7 ) Wingspan 12–14 mm. Head ochre, nearly white around eye. Labial palp white on inner side, brown on outer side; 3rd article about half length of 2nd. Antennal flagellum annulated with dark brown and white; scape brown with prominent tuft of long, erect, brown scales. Thorax white. Tegula ochre with some white on outer margin. Forewing ochre with dull white streaks: costal streak thin, gradually widening toward fringe; fringe white in proximal 1/3, brown distally; discal streak at dorsal edge of cell, widened medially and interrupted before termen; dorsal streak along fold; a thin white line bordering dorsal margin to terminal fringe; fringe pale brown. Hindwing brown with brown fringe. Abdomen greyish brown.
♀ ( Figs. 2, 7 View FIGURES 1 – 7 ) Wingspan 10–11.5 mm. Forewing paler than in male, pale streaks shiny white; pale costal streak wider distally and extended to costal fringe; medial white streak nearly straight, shorter and thinner than in male.
Male genitalia ( Figs. 8–10 View FIGURES 8 – 12 , 13–14 View FIGURES 13 – 16 ): Spinose knob of gnathos rounded. Tegumen short, squat, with short pedunculi. Transtilla thin, elongate, oblique. Valvula large in proportion to sacculus, well delineated with rounded ventral edge. Cucullus narrowly elongate, slightly wider apically. Sacculus subrectangular, ventral edge slightly arched, lateral edge almost straight or slightly curved, with ventro-caudal angle rounded. Phallotheca tubular, distally narrowed, dorsal side more sclerotized. Cornuti about 20 in number, small and grouped into thin, somewhat inconspicuous band.
Female genitalia ( Figs 12 View FIGURES 8 – 12 , 17, 20 View FIGURES 17 – 22 ): Papillae anales transversely broad, oval. Posterior apophysis about 3x longer than anterior one. Sterigma subtrapezoid in outline, wider than long, irregularly melanized; distal edge convex around ostium bursae and with elongate, stout spines. Ostium bursae cup-shaped, squarish. Colliculum shaped like very elongate funnel with sides more sclerotized than middle, with thin dark midline. Ductus bursae with distal (caudal) section almost transparent, with thick, dark midline and with short finely spinulose section just anterad of colliculum; middle section papillate around inception of ductus seminalis; anterior section to corpus bursae membranous. Corpus bursae rounded; signum large, thorn-like with strong, curved hook and relatively narrow base.
Abdominal apodemes ( Fig. 11 View FIGURES 8 – 12 ): Latero-posterior strut about 2x longer than anterior strut. Transverse strut thick, distal edge more sclerotized. Spine patches of tergum 3 with 12–16 spines.
Larval case ( Figs. 24–26 View FIGURES 23 – 27 ): similar to those of C. pyrrhulipennella and C. pulchripennella ; made up entirely of lustrous black silk; 7–8 mm in length, elongate, subcylindrical with a slight curvature, laterally compressed, with the oral end narrower and oval at a 10–30˚ angle; anal end rounded with a bivalved opening; surface of case with obvious growth lines, and about the middle on the dorsal side is a small brown patch which is the remnant of the early case; the ventral side has a weakly developed keel.
Biology. The larval host plant is Erica carnea L. ( Ericaceae ). The early larval instars have not been observed. However, it is likely that the life cycle is similar to that of C. pyrrhulipennella which is described in Emmet et al. (1996). Partly grown larvae were found (in Lavant, Osttirol) in the first half of May mining new terminal leaves of the host plant. At the end of larval growth, the final case is attached on a twig in a sun-exposed location, with the anal end turned upward. The flight period occurs between June 20 and the middle of July, with variations at either end depending on elevation and yearly seasonal variation. The species has been collected at elevations between 250 m and 1600 m.
Distribution. Northern Italy (Trentino-Alto Adige and Friuli); southern Austria (Osttirol); Slovenia. The westernmost and southernmost locality is Pietramurata in Trentino north of Lago di Garda, whereas the northernmost locality is Lavant in East Tyrol, and the easternmost is Mount Triglav in Slovenia. Records from Italy were previously referred to as C. pyrrhulipennella ( Baldizzone 1987, 1996; Baldizzone & van der Wolf 2004) but it now appears that that species does not occur in Italy.
DNA barcode analysis. Tissue samples (dried legs) were shipped to the Canadian Centre for DNA Barcoding in Guelph for DNA extraction, amplification, and sequencing following standard protocols (deWaard et al. 2008). Barcoding efforts included the holotype and 6 paratypes of the new species as well as representatives of C. pulchripennella and C. pyrrhulipennella , including one record of the latter obtained from GenBank. The Barcode Identification Numbers (BINs) ( Ratnasingham & Hebert 2013) in BOLD are used as registry designations for barcode clusters. Neighbor-joining trees and genetic distances were calculated with the Taxon ID Tree and Distance Analysis tools available in BOLD using the Kimura two-parameter (K2P) model of base substitution and Kalign sequence alignment. Details of the barcoded specimens and their photographs are available through the following dataset (http://dx.doi.org/10.5883/DS-COLERICA). The same DOI provides access to the sequence records, trace files, and primer sequences used for PCR amplification, together with GenBank accession numbers. Twenty-eight specimens were analyzed, resulting in 658 bp, full-length barcode fragments for 21 specimens. A sequence of 617 bp with 75 ambiguous positions was obtained for a specimen of C. pyrrhulipennella (CNCLEP00068773), which was excluded from the NJ analysis. Attempt to barcode the holotype of C. pulchripennella failed due to contamination problems. These shortcomings did not affect the overall results of the barcode analysis.
Genetic differences within this group of closely related species are relatively small ranging from 1.5–2.0%, but cluster separation is sufficiently cohesive to result in the attribution of different BINs ( Table 1 View TABLE 1 , Fig. 28 View FIGURE 28 ). Despite the small barcode gaps, genetic differences are congruent with the differences observed in genitalia morphology, larval host plant, as well as the allopatric distribution of C. ericarnella relative to its nearest neighbor C. pyrrhulipennella . Coleophora ericarnella is restricted to the south-eastern part of the Alps, whereas C. pyrrhulipennella is relatively widespread in Europe north of the Alps but absent from Italy. The more distinct C. pulchripennella has a Mediterranean distribution and its larval host is Erica arborea .
All three species form a compact group that is genetically distinct and divergent from the remainder of Palearctic Coleophora when their barcodes are compared against the entire BOLD library, which includes barcodes from over 1200 species of Coleophora . DNA barcoding has been used successfully to elucidate cryptic species diversity in several genera of Microlepidoptera where morphological characters, particularly in the genitalia, are subtle ( Huemer & Hebert 2011; Baldizzone & Landry 2011; Huemer & Mutanen 2012; Baldizzone et al. 2014; Huemer & Timossi 2014; Huemer et al. 2014; Huemer & Mutanen 2015). Coleophora ericarnella represents another example of suspected cryptic species where DNA barcodes provides additional support for the status of distinct species.
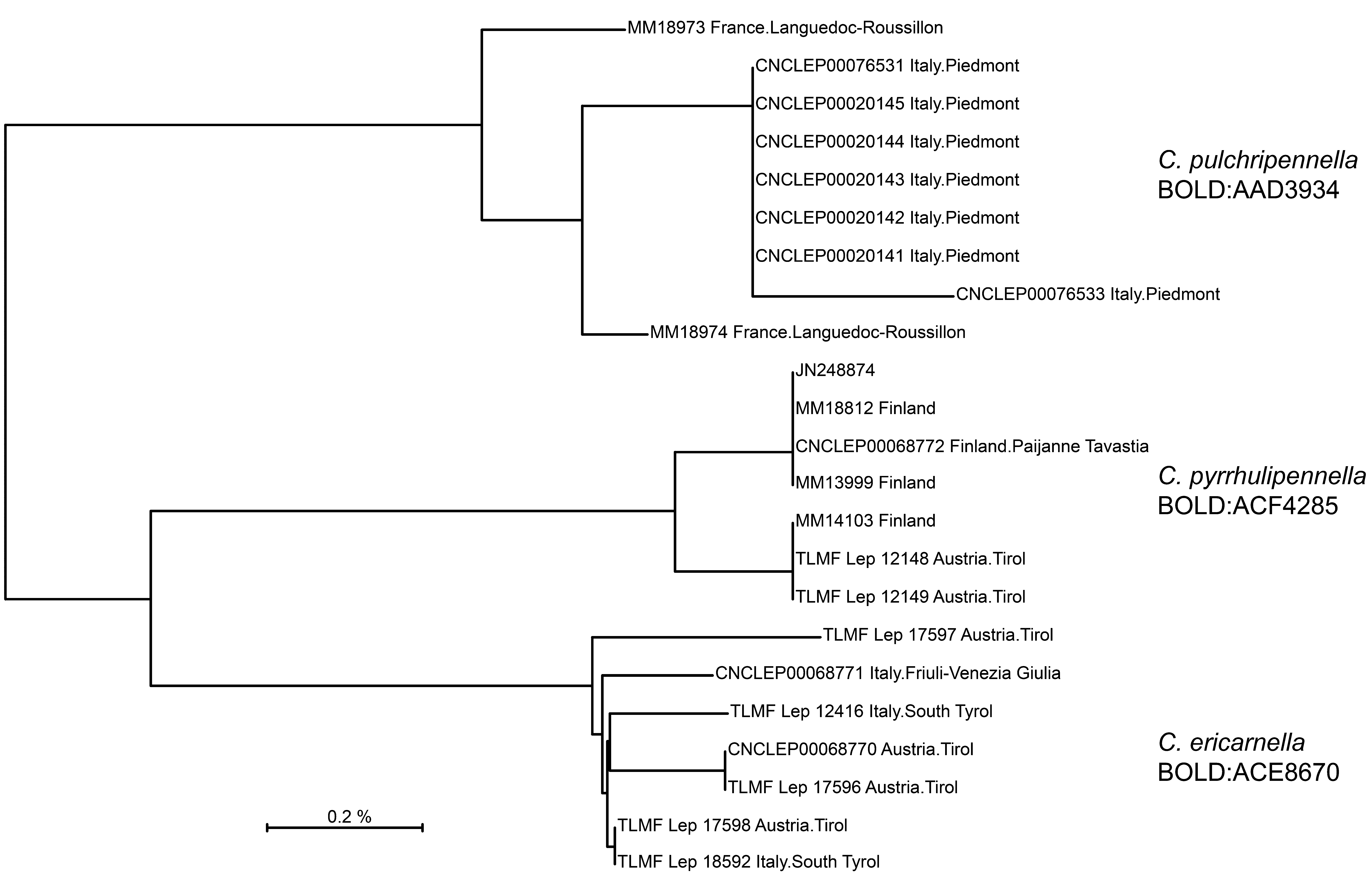

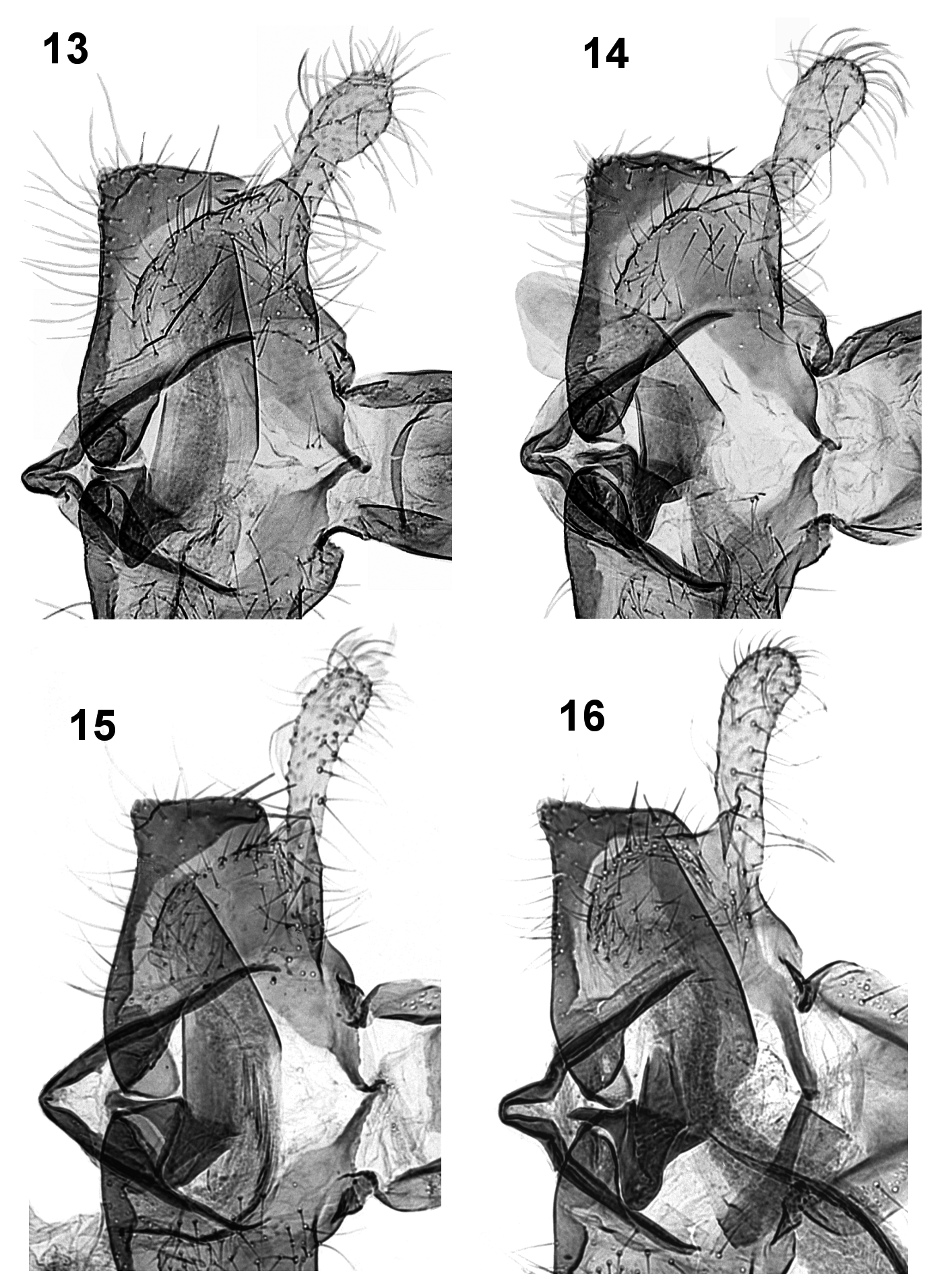
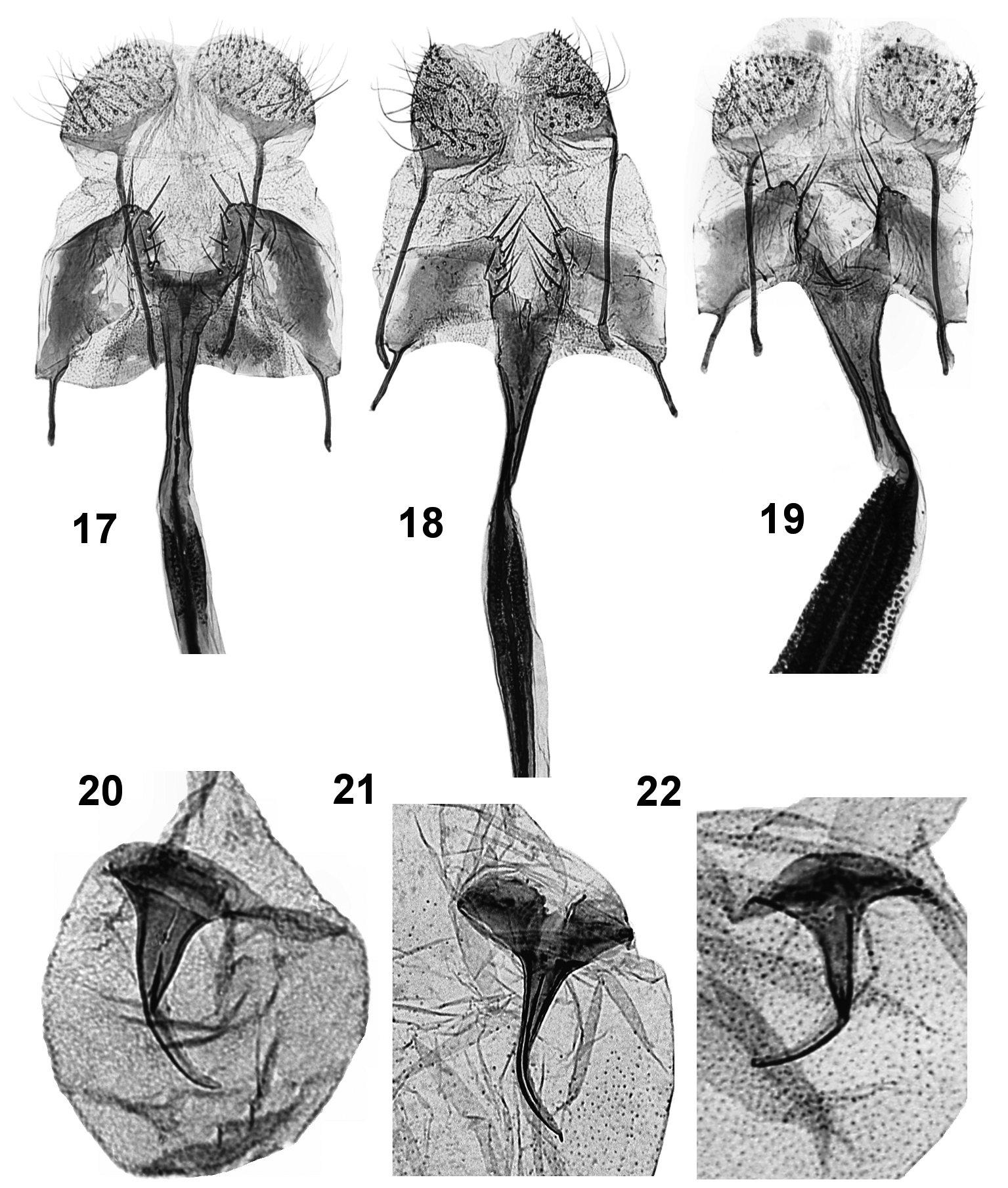
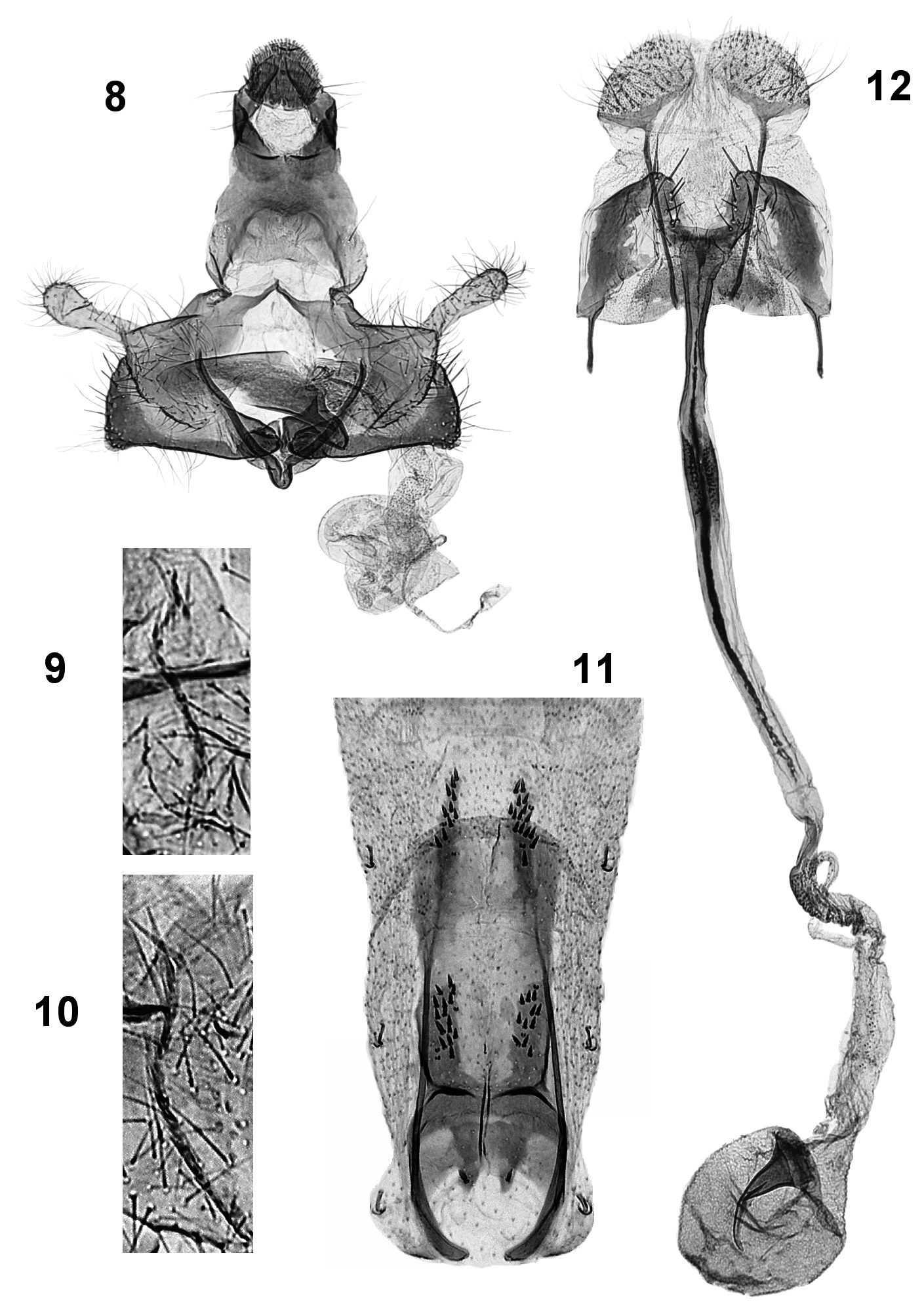

TABLE 1. Percentage of divergence in DNA barcode (cytochrome c oxidase I gene) sequences among three species of Coleophora of the pyrrhulipennella complex. “ BOLD: ABC 1234 ” = Barcode Index Number. Diagonal cells = intra-specific distances; cells below diagonal = mean inter-specific distances; S. E. = standard error.
| C. ericarnella | C. pyrrhulipennella | C. pulchripennella | |
|---|---|---|---|
| C. ericarnella BOLD:ACE8670 (n=7) | Max = 0.46 Average = 0.25 S.E. = 0.03 | ||
| C. pyrrhulipennella BOLD:ACF4285 (n=7) | Min = 1.37 Average = 1.52 S.E. = 0.02 | Max = 0.31 Average = 0.17 S.E. = 0.04 | |
| C. pulchripennella BOLD:AAD3934 (n=9) | Min = 1.54 Average = 2.09 S.E. = 0.07 | Min = 1.70 Average = 2.04 S.E. = 0.03 | Max = 0.79 Average = 0.23 S.E. = 0.04 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |