
Phorbas topsenti, Vacelet, Jean & Perez, Thierry, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.184048 |
|
DOI |
https://doi.org/10.5281/zenodo.5614632 |
|
persistent identifier |
https://treatment.plazi.org/id/03FD2132-FFB9-FFDD-888B-CDE5FAA9FE92 |
|
treatment provided by |
Plazi |
|
scientific name |
Phorbas topsenti |
| status |
sp. nov. |
Phorbas topsenti View in CoL sp. nov.
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Restricted synonymy. Anchinoe paupertas: Topsent, 1934: 25 ; Topsent, 1936: 43; Boury-Esnault, 1971: 318; Pansini, 1987: 166. Phorbas paupertas: Maldonado, 1993: 244 ; Carballo, 1994: 148.
Holotype. MNHN DJV 113, Marseille, Anse des Cuivres near Station Marine d’Endoume, 43°16'47.46"N, 5°20'57.22"E, 7 m, 21 July 2004.
Paratype 1. MNHN DJV 114, Marseille, Anse des Cuivres near Station Marine d’Endoume, 43°16'47.46"N, 5°20'57.22"E, 7 m, 21 July 2004.
Paratype 2. MNHN DJV 115, Lebanon, Beirut harbour, 33°54.36’N, 35°30.98’E, 15 m, 9 July 2003.
Paratype 3 MNHN DJV 116, Tunisia (Kerkennah Islands), 4 m, 16 December 1987.
Other material examined. Numerous specimens from Marseille, Tunisia and Lebanon.
Description. Thickly encrusting to massive ( Fig. 1 View FIGURE 1 ), thickness varying from a few millimeters to 25 mm, covering areas up to 100 cm ². Surface smooth, entirely covered by pore-sieves, 1.5–2 mm in diameter. Oscules scattered, 2–3 mm in diameter in preserved specimens. Consistency fleshy, rather soft. Colour deep red in live specimens, cream in alcohol.
Skeleton. Dermal skeleton of dispersed strongyles, mostly tangential to the surface between the poresieves, and arranged in a dense palisade around the pore-sieves ( Fig. 2 View FIGURE 2 A). Choanosomal skeleton ( Fig. 2 View FIGURE 2 B) consisting of plumose ascending fibres, 80 µm in maximum thickness, with strongyles and acanthostyles I included in a feebly developed spongin, echinated by acanthostyles II and a few acanthostyles I. Isochelae present everywhere in the sponge, more numerous in the dermal membrane.
Spicules ( Fig. 3 View FIGURE 3 A–F, Tables 1 View TABLE 1 and 2 View TABLE 2 ). Strongyles or subtylostrongyles, straight, frequently with slightly enlarged ovoid ends. Acanthostyles I, slightly curved, with a few short, generally blunt spines near the head.
Acanthostyles II, straight, slightly conical, entirely spined, spines sharply pointed, curved toward the head along the shaft. Isochelae, arcuate, with shaft 3–3.3 µm in diameter, oval alae 8 µm long, central tooth wellformed, rudimentary fimbriae barely visible on some spicules.
These spicules show rather low variations, according to the paratypes, to other specimens examined and to the literature (tables 1 and 2). The identification as Anchinoe paupertas by Pulitzer-Finali 1983 for specimens from the Italian coasts which are thinly encrusting and with no reported isochelae need to be checked.
Histology and cytology ( Fig. 4 View FIGURE 4 ). Choanocyte chambers ( Fig. 4 View FIGURE 4 A, B) spherical, 23–26 µm in diameter, with small choanocytes. Two types of cells with inclusions present. Vacuolar cells ( Fig. 4 View FIGURE 4 C, D), numerous, generally ovoid with irregular outline, up to 14 x 6 µm, with a small, anucleate nucleus; cytoplasm more or less reduced between clear vacuoles with a sinuous outline. Vacuoles up to 3 µm in diameter, occupying most of the cell volume. Spherulous cells ( Fig. 4 View FIGURE 4 E), subspherical to elongate, up to 16 x 6 µm; nucleus with a central area containing a large crystal-like inclusion; spherules dense, homogeneous, 1–2 µm in diameter. Rather dense fascicles of collagen fibrils ( Fig. 4 View FIGURE 4 F) present in places. Two morphotypes of symbiotic bacteria present in the mesohyl although not abundant, rod-like bacteria ( Fig. 4 View FIGURE 4 F) and remarkable spiral-shaped bacteria ( Fig. 4 View FIGURE 4 G).
Reproduction. A specimen from Lebanon (collected on 05/06/2000: 33a) contained numerous embryos, spherical (150–200 µm in diameter) in the early stages and ovoid, up to 400 x 250 µm in the most mature stage where they contain thin strongyles (80–110 x 0.5 µm) dispersed without order, and incompletely developed isochelae. Spermatogenesis has been observed in a specimen collected in Marseille (30/05/1984) (see De Vos et al. 1991).
Symbiosis. The sponge is frequently associated with the amphipod Tritaeta gibbosa (Bate, 1862) .
Ecology. Common in well lit or semi-shaded areas, between 2 to 50 m.
Distribution. Mediterranean ( Tunisia, French, Italian and Spanish coasts, Gibraltar, Lebanon). The sponge is recorded from the Strait of Gibraltar ( Carballo 1994), but the records as Anchinoe , Phorbas or Hymedesmia paupertas from outside the Mediterranean refer to Hymedesmia paupertas (for instance Cristobo et al. 1998 in Galicia).
Etymology. The species is dedicated to Émile Topsent who first recognized this sponge in the Mediterranean.
Remarks. This sponge was first described in the Mediterranean by Topsent (1934), who recorded it from Tunisia and from the Mediterranean French coasts. Topsent wrongly identified the sponge as Anchinoe (now Phorbas ) paupertas , described from the north European coasts by Bowerbank (1866) as Hymeniacidon paupertas , which at that time was poorly defined and which is a Hymedesmia . Recent revisions have shown that Hymedesmia paupertas (Bowerbank) is a thinly encrusting, blue or green sponge from the North East Atlantic, with an hymedesmioid skeleton, which is clearly different from the red Mediterranean sponge (van Soest et al. 2000). We thus propose a new name for the Mediterranean sponge, which has often been recorded as Phorbas paupertas . The ‘true’ Hymedesmia paupertas has apparently not been observed in the Mediterranean.
Phorbas topsenti View in CoL sp.nov. closely resembles by its field characters two sponges of the same family, Phorbas fictitius (Bowerbank, 1866) View in CoL and Hamigera View in CoL hamigera (Schmidt, 1862) View in CoL . The three sponges are clearly distinct by their spiculation, the major differences being the presence of tornotes instead of strongyles in P. fictitius View in CoL , and the absence of true acanthostyles in H. hamigera View in CoL ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ). However, they are extremely difficult to distinguish in the field. Phorbas fictitius View in CoL appears to have a more variable colour, varying from deep red to grey and pale orange pink in the NE Atlantic (van Soest et al. 2000). However, in the NW Mediterranean, P. fictitius View in CoL appears always as red, possibly a little more vivid than P. topsenti View in CoL sp.nov. The three species have the same ecological distribution, but are generally not found together in the same area. In our experience on the French coasts of the NW Mediterranean, the species most commonly found in the Marseille area is P. topsenti View in CoL sp.nov., whereas P. fictitius View in CoL is more common in Port-Cros and H. hamigera View in CoL in Banyuls. The present cytological study shows that P. topsenti View in CoL sp.nov. and H. hamigera View in CoL are closely related, having similar vacuolar and spherulous cells, including crystal-like nuclear inclusion, and similar spiral-shaped bacteria ( Boury-Esnault 1972, 1976). These bacteria have been rarely observed in other sponges, but resemble to a greater or lesser extent those described in Grantia compressa ( Vacelet & Donadey 1977) View in CoL , in Clathrina cerebrum ( Burlando et al. 1988) View in CoL , and in Halisarca dujardini ( Ereskovsky et al. 2005) , which have no relationship with these poecilosclerids.

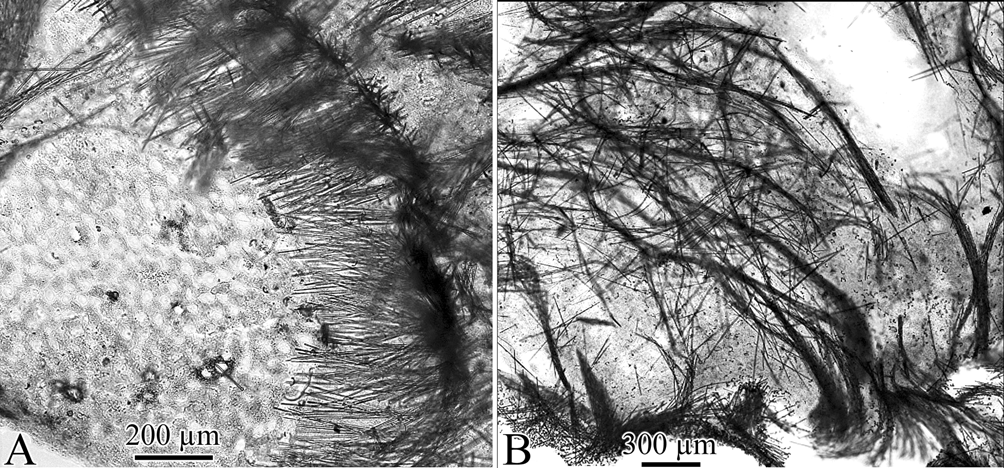

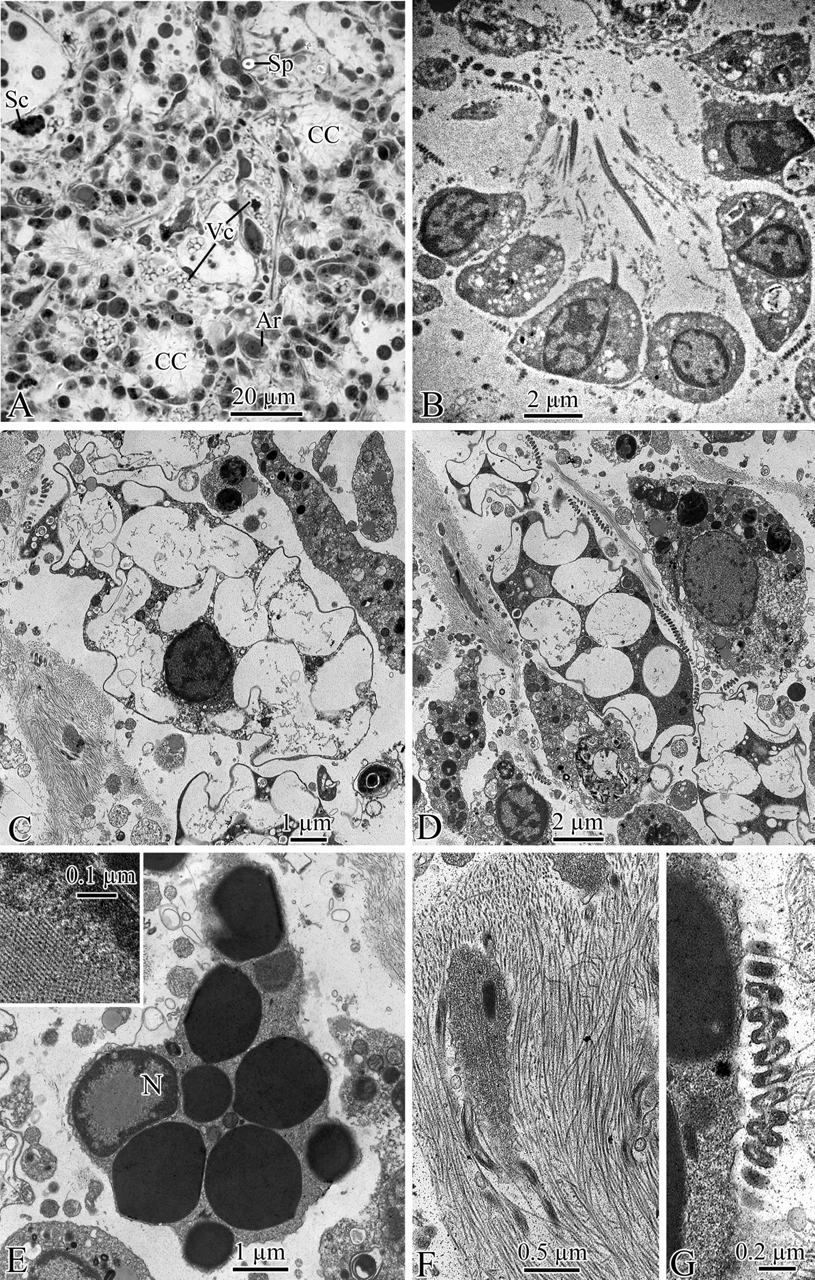


TABLE 1. Phorbas topsenti sp. nov. Spicule size (µm) in the type material.
| Strongyles | Acanthostyles I | Acanthostyles II | Isochelae | |
|---|---|---|---|---|
| Holotype, Marseille | 279.5 ± 13.2 (260–305) x 4.0 ± 0.8 (2.75–5.25) | 329.3 ± 16.9 (300–360) x 6.2 ± 0.9 (4.5–7.5) | 93.8 ± 11.5 (70–110) x 3.9 ± 0.8 (2.5–5.25) | 22.3 ± 1.6 (20–25) |
| Paratype 1, Marseille | 262.7 ± 13.0 (230–282) x 3.9 ± 1.0 (2.5–6) | 329.4 ± 19.99 (285– 360) x 5.88 ± 0 78 (4–7) | 87.52 ± 5.61 (70–100) x 4.06 ± 1.06 (2.5–6) | 21.1 ± 1.1 (18.5–22.5) |
| Paratype 2 (Lebanon) | 286.3 ± 16.4 (242–320) x 3.8 ± 0.9 (2.5–5) | 393.2 ± 19.63 (350– 430) x 4.94 ±1.04 (3–6) | 97.1 ± 4.45 (80–105) x 3.98 ± 0.84 (2–5) | 25 ± 0.5 (24–26.5) |
| Paratype 3 (Tunisia) | 243.6 ± 16.3 (210–270 x 4.4 ± 0.64 (2.5–5) | 289.2 ± 14.68 (260– 320) x 6.6 ± 1.26 (4–8) | 92.2 ± 5.10 (78–102) x 5.2 ± 0.6 (4–6) | 22.2 ± 1.1 (20.2–24.2) |
TABLE 2. Spicule size (µm) of Phorbas topsenti in the literature.
| Strongyles | AcanthostylesI | AcanthostylesII | Isochelae |
|---|---|---|---|
| Topsent 1934 Tunisia (La Skira) 275 x 4 | 350 x 6 | 100 x 4–5 | 25 |
| Topsent 1934 Tunisia (Adjim) 275–285 x 3–4 | 325–335 x 6 | 100 x 4–5 | 21–24 |
| Topsent 1934 La Ciotat 300–330 x 4 | 330–375 x 7 | ? x 5 | 24–25 |
| Topsent 1934 Monaco 300–325 x 4 | 375–400 x 8 | 135 x 6 | 32 |
| Topsent 1934 Monaco 275 x 4 | 310–350 | ? | 32 |
| Topsent 1934 Banyuls 200–240 | 250–350 x 7 | ? x 5 | 19–21 |
| Maldonado 1993 Alboran 200–323 x 2–3 | 330–550 x 8–10 | 80–160 x 6–7 | 23–32 |
| Carballo 1994 Gibraltar 219–280 x 1.9–4.2 | 300–370 x 6–9 | 95–125 x 5–7 | 24–33 |
| Boury-Esnault 1991 Banyuls 248–317 | 274–446 | 79–140 | 23.4–30.6 |
| Pansini 1987 Straight of Sicily 235–280 x 3–5 | 270–320 x 4–6 | Absent | 19–24 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phorbas topsenti
| Vacelet, Jean & Perez, Thierry 2008 |
Halisarca dujardini (
| Ereskovsky et al. 2005 |
Clathrina cerebrum (
| Burlando et al. 1988 |
Grantia compressa (
| Vacelet & Donadey 1977 |
Phorbas fictitius
| Bowerbank 1866 |
hamigera
| Schmidt 1862 |