
Hassar affinis ( Steindachner, 1881 )
|
publication ID |
https://doi.org/ 10.1590/S1679-62252011000300006 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE1801-FFC3-FF8D-D65E-F8A2DD46F96B |
|
treatment provided by |
Carolina |
|
scientific name |
Hassar affinis ( Steindachner, 1881 ) |
| status |
|
Hassar affinis ( Steindachner, 1881) View in CoL
Figs. 2 View Fig and 3 View Fig
Oxydoras affinis Steindachner, 1881: 107 View in CoL , pl. 1, figs. 1, 1a [type locality: “ Rio Puty ” (= rio Poti, Piauí, Brazil)].
Hemidoras affinis .— Eigenmann & Eigenmann, 1888: 158 [change of generic status].— Eigenmann & Eigenmann, 1890: 258 [description based on six specimens from rio Poti, probably collected with holotype].— Eigenmann & Eigenmann, 1891: 33 [literature compilation].— Eigenmann, 1910: 394 [literature compilation].— Steindachner, 1915: 73 [description, Itapicuru (=Itapecuru) at Caxias, Paranaíba (=Parnaíba) at Engenho d’Água].
Hassar affinis View in CoL .— Kindle, 1895 [change of generic status, identification key].— Miranda Ribeiro, 1911: 188 [identification key; translation to portuguese of the description given by Eigenmann & Eigenmann, 1890].— Eigenmann, 1925: 355, pl. 27, fig. 4 [identification key, literature compilation, illustration].— Gosline, 1945: 22 [literature compilation].— Van der Stigchel, 1946: 95 [description based on a single specimen from Rio Puty (=rio Poti)].— Menezes, 1949: 93 [feeding habit, common name, rio Poti Velho, rio Parnaíba at Terezina, Lagoa de Nazaré at Floriano, rio Piauí at Poço do Salão, São João do Piauí].— Fowler, 1951: 491 [literature compilation, see notes on Distribution].— Eschmeyer, 1998: 49 [literature compilation].—Sabaj & Ferraris, 2003: 461 [literature compilation].— Ferraris, 2007: 171 [literature compilation].— Birindelli et al., 2009: 276 [gas bladder morphology].
Hassar woodi Fowler, 1941a: 139 View in CoL , figs. 35, 36, 37 [type locality: Rio Parnahyba, Therezina, Piauhy (= rio Parnaíba, Teresina, Piauí)].— Gosline, 1945: 23 [literature compilation].— Fowler, 1951: 493 [literature compilation].— Eschmeyer, 1998: 1796 [literature compilation].
Hassar wilderi View in CoL .— Soares, 2005: 90 [description, illustration, rio Mearim].
Type-specimens. ANSP 69392 About ANSP (1, 201.6 mm SL, holotype of Hassar woodi View in CoL ), rio Parnaíba , Teresina, PI, Brazil , 1936, R. von Ihering. ANSP 69396 About ANSP (1, 76.0 mm SL, paratype of Hassar iheringi View in CoL ), Ceará [presumably purchased in the fish market at Fortaleza, CE], Brazil , 1937, R. von Ihering. NMW 45394 (1, 123.0 mm SL, holotype of Hassar affinis View in CoL ), Rio Puty [=rio Poti, 5°5’S 42°49’W, Teresina, PI, Dec 1865, O. St. John], Brazil GoogleMaps .
Non-type specimens. Itapecuru basin ( Brazil): MZUSP 104559 View Materials (3, 113.0-128.0 mm SL), rio Itapecuru , 3°31’39’’S 44°24’19’’W, Itapecuru-Mirim, MA, 2006, M. C. Barros et al. Parnaíba basin ( Brazil): MCZ 7200 (4, 109.0- 197.6 mm SL); BMNH 89.11 .14.46 (1, 120.7 mm SL); collected with holotype. MZUSP 5123 View Materials (2, 127.2- 129.1 mm SL), rio Parnaíba , Teresina, PI. MZUSP 36454 View Materials (2, 126.6- 140.3 mm SL), Teresina, PI. MZUSP 36602 View Materials (1, 110.0 mm SL), Teresina, PI. MZUSP 37888 View Materials (2, 74.8-101.9 mm SL), rio Sambito , São Félix, PI. MZUSP 74914 View Materials (3, 102.6- 104.3 mm SL), Teresina, PI. MZUSP 74890 View Materials (1 c&s, 5, 86.8-96.8 mm SL), rio Poti , Teresina, PI, Nov 2001, M. C. C. de Pinna. MZUSP 74919 View Materials (1, 97.7 mm SL), rio Sorubim , Campo Maior, PI. MZUSP 89935 View Materials (1 c&s, 11, 105.6- 149.2 mm SL), Island at Lagoa de Parnaguá , 10°14’41”S 44°38’50”W, Parnaguá, PI, Apr 2001, Expedição Jalapão [O. T. Oyakawa et al]. Pindaré-Mearim basin ( Brazil): MZUSP 5067 View Materials (1, 86.7 mm SL), rio Grajaú , Grajaú, MA. MZUSP 43604 View Materials (4, 141.1-157.0 mm SL), Lago dos Viana, c. 3°13’S 45°10’W, MA, Apr 1984, Moura. MZUSP 79833 View Materials (2, 135.2- 141.1 mm SL), rio Maracu , MA, Nov 1984. MZUSP 87493 View Materials (2, 122.0- 146.6 mm SL), rio Balsas , 7°32’8”S 46°2’16”W, Balsas, MA, Mar 2005, A.Akama & E. Baena. Turiaçu basin ( Brazil): CPDOL ( UFMA) 98186 (12, 90.7-126.1 mm SL), rio Turiaçu basin, Oct 1999, N. Piorski & L. Pereira. MPEG 3014 View Materials (1, 134.0 mm SL), rio Turiaçu, Lagoa Guajá, near Posto Indígena Guajá, MA GoogleMaps .
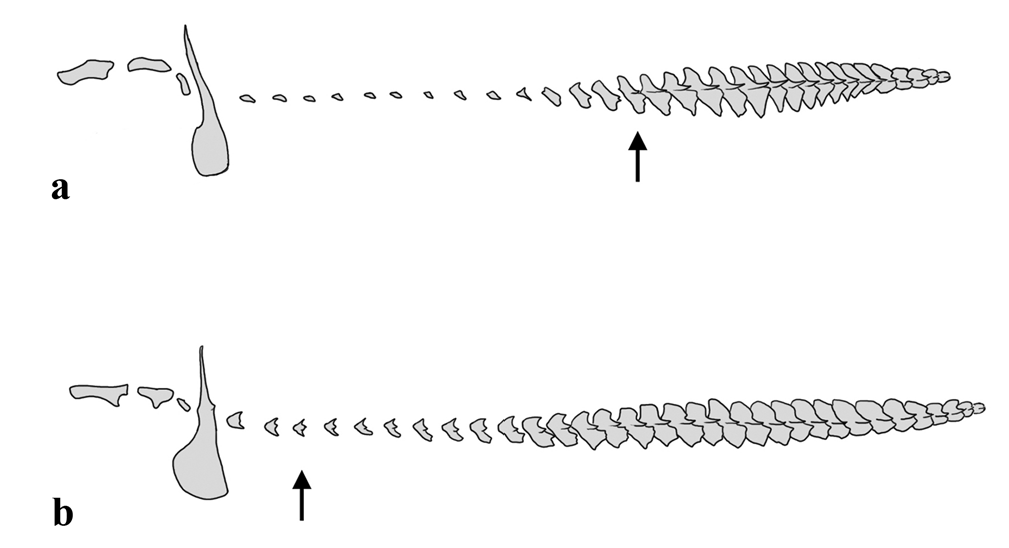
Diagnosis. Hassar affinis is diagnosed among congeners by having the distal tip of the anterior branched dorsal-fin rays and membranes darkened (vs. distal tip of anterior branched dorsal-fin rays and membranes pale). Hassar affinis is distinguished from H. orestis and H. wilderi by having two rounded diverticula restricted to each side of anterior chamber of gas bladder (rarely one extra pair on posterior chambers) (vs. many well-branched diverticula along margins of entire gas bladder) ( Fig. 4 View Fig ); and gas bladder rounded posteriorly (vs. gas bladder triangular posteriorly, each posterior chamber extended into a short terminal diverticulum sharing medial septum with its pair). Hassar affinis is further distinguished from H. orestis by having 12 th through 17 th, modally 13 th, midlateral scute as the anteriormost with median thorn (vs. 1 st through 8 th, modally 3 rd) ( Fig. 5 View Fig ), 10 th midlateral scute 3.1-5.1%, mean 4.1%, of relative body depth (vs. 6.2-18.0%, mean 12.9%, of relative body depth), and tip of upper caudal-fin lobe not darkened (vs. usually darkened). Hassar affinis is distinguished from H. gabiru by having lateral diverticula on gas bladder rounded (rarely branched in large specimens) (vs. lateral diverticula finger-like, weakly-branched in large specimens).
Description. Morphometric data are summarized in Tables 1 and 2; type specimens illustrated in Fig. 2 View Fig ; additional specimens in Fig. 3 View Fig . Largest specimen examined 201.6 mm SL (ANSP 69392, holotype of Hassar woodi ); reported to 213 mm SL ( Menezes, 1949). Dorsal profile of head rising moderately, evenly (usually in smaller specimens) or strongly convex (especially in larger specimens) from snout tip to anterior margin of orbit, and relatively straight from latter point to dorsal-fin spine. Dorsal profile of body descending gradually, approximately straight from dorsal-fin spine to caudal peduncle. Ventral contour shallowly concave from snout tip to pectoral girdle, and slightly convex from latter point to caudal peduncle. Caudal peduncle short with shallow hourglass shape in lateral view.
Body elongate with prominent conical snout. Mouth subterminal,each premaxilla bearing small patch of approximately 5 to 10 acicular teeth, and each dentary with approximately 10 to 20 acicular teeth. Oval orbit with weakly developed adipose eyelid in juveniles and adults, extended slightly beyond anterior and posterior limits of eye. Eyes positioned about half way between tip of snout and dorsal-fin origin.
Three pairs of barbels (maxillary, inner and outer mental), all fimbriate. Maxillary barbel usually reaching base of first pectoral-fin ray; with 7 to 16 (mode 11, n=47) fimbriae along ventrolateral face. Inner and outer mentonian barbels of approximately equal size, covered with many rounded papillae, and falling short of ventralmost opening of gill slit.
Gill rakers small on first gill arch, absent on remaining arches.Accessory branchial lamellae on inner face of first gill arch well developed in approximately ten rows from insertion of rakers to origin of branchial filaments (but not contacting the latter); accessory lamellae gradually reduced on remaining (second to fifth) gill arches.
Lateral-line tubules ossified, forming row of 31 to 34 (mode 32, n=17) midlateral scutes beginning with infranuchal. Three tympanal scutes, inconspicuous, usually without emergent thorn. Infranuchal scute with dorsal wing extremely thin and ventral wing dilated, expanded anteriorly, connected to posterior cleithral process; scute usually without medial thorn. Postinfranuchal scutes reduced anteriorly, non-overlapping; each with posterior margin bicuspid (without medial thorn) or tricuspid (including medial thorn), latter condition usually starting at 13 th scute (range 12 th through 17 th, n=19); medial thorn and dorsal and ventral wings gradually increasing in size posteriorly; scutes with serrated posterior margins and overlapping on the last third of the body.
Dorsal-fin II,6 (n=52), triangular with distal margin approximately straight, vertical when erected. Dorsal-fin spine slightly compressed and curved, with relatively small antrorse serrations along anterior margin (serrations reduced or absent on distal third); slightly larger retrorse serrations along posterior margin (serrations absent on proximal portion). Pectoral fin modally I,8, range I,7-9 (n=52); distal margin straight, oblique relative to body axis. Pectoral-fin spine slightly depressed and curved, with antrorse serrations along anterior margin (serrations absent on distal third); slightly larger retrorse serrations along posterior margin (serrations larger distally). Pelvic fin i,6 (n=52); distal margin rounded. Anal fin modally iv,8, range iii-v,7-9 (n=51); subtriangular with scarcely rounded distal margin. Adipose fin relatively small, teardrop-shaped, with rounded free posterior margin. Caudal fin i,7/8,i (n=52, rarely i,7/7,i or i,7/ 9,i), distinctly forked, with lobes approximately equal in size.
Gas bladder ( Fig. 4 View Fig ) moderately large, cordiform. Gas bladder walls entirely smooth in small specimens (up to 50 mm SL). Large specimens have only two short rounded diverticula on each side of anterior chamber; in the largest specimen examined (ANSP 69392, 201.6 mm SL) a third pair of diverticula are present at middle of lateral margin of posterior chambers; lateral diverticula rarely branched (only in large specimens). Gas bladder rounded posteriorly, not extended into terminal diverticula.
Osteology. Osteology generally similar to that described for Hassar orestis , excepting differences as follows. Total vertebrae 34 (n=1) or 36 (n=1), vertebrae 6-13 bearing ribs. Eight (n=2) dorsal-fin pterygiophores, 11 (n=2) pelvic-fin pterygiophores; 12 (n=1) or 13 (n=1) dorsal and 12 (n=1) ventral caudal-fin procurrent rays. All examined specimens with hypural fusion pattern PH; HY 1+2; HY 3+4; HY 5, and with complete ossification of sutural joint at junction of sphenotic, parieto-supraoccipital and frontal.
Coloration. In alcohol, head and body tan to brown, or grey, countershaded. Faint dark stripe from anterior margin of upper lip to anterior margin of eye. First two (or rarely three) branched dorsal-fin rays and membranes distally darkened (distalmost fifth to third of fin-ray length), dark pigment extending to tips of rays and membranes (except in some large specimens with dorsal fin uniformely tan). Tip of upper caudal-fin lobe not conspicuously darkened. Soares (2005) noted the upper sides and flanks yellowish-gray and ventral surfaces white in a live specimen of Hassar affinis .
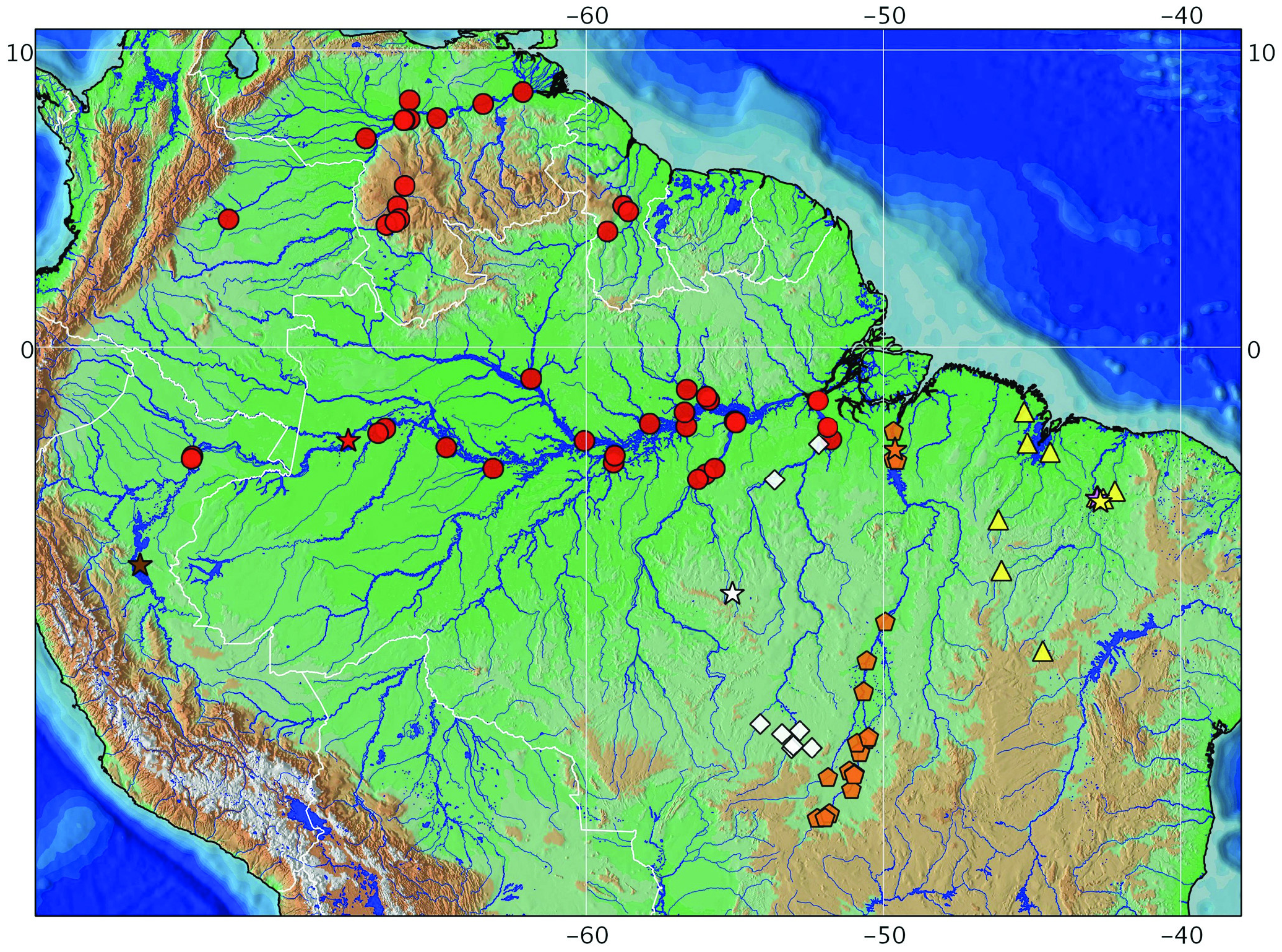
Distribution. Hassar affinis occurs in northeastern Brazil, including Turiaçu, Itapecuru, Pindaré-Mearim, and Parnaíba basins ( Fig. 6 View Fig ). Only one other doradid species occurs in those northeastern Brazilian drainages, Platydoras brachylecis , which is also endemic to the area ( Piorski et al., 2008). Eigenmann & Fisher (1917) mistakenly identified a specimen from San Joaquin, in Bolivia, as Hemidoras affinis (= Hassar affinis ). Fowler (1951) mentioned that Hassar affinis occurs in “Amazônia, Bolívia, Rio Paranaíba, Rio Itapicuru na Bahia”; but the species actually occurs in rio Parnaíba and rio Itapecuru (not Paranaíba, nor Itapicuru in Bahia).
Ecology. Menezes (1949) examined the diet of 25 specimens (48-180 mm SL) collected in lentic and lotic environments of the rio Parnaíba, Piauí, and observed that H. affinis feeds primarily on insects and, to a lesser extent, on crustaceans and plant residue. Fish remains were found in the intestine of a large examined specimen (213 mm SL). According to Soares (2005) Hassar affinis inhabits igarapés, lakes and rivers, and has an omnivorous diet.
Etymology. Steindachner (1881) described Oxydoras affinis (= Hassar affinis ) as a variation of O. orestis (= H. orestis ), a species he considered closely related or adjacent to.
Remarks. Steindachner (1881) distinguished Hassar affinis by having 18 to 20 midlateral scutes with thorns (vs. more in H. orestis ), and a “slightly darker coloration on the superior marginal portion of the first rays of [dorsal] fin” (vs. a “sharply defined black mark” on the dorsal fin in H. orestis ). Both characters are corroborated herein as consistent differences between the two species.
Fowler (1941a) reported a single juvenile specimen (ANSP 69396, 76 mm SL) as collected in Fortaleza (Ceará). However, since no large drainage exists between Fortaleza and rio Parnaíba, Sabaj (2002) suggested that that specimen was probably obtained in a fish market, making it unfeasible to substantiate its origin.



| NMW |
Naturhistorisches Museum, Wien |
| UFMA |
Universidade Federal do Maranhão |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hassar affinis ( Steindachner, 1881 )
| Birindelli, José L. O., Fayal, Danielle F. & Wosiacki, Wolmar B. 2011 |
Hassar wilderi
| Soares, E 2005: 90 |
Hassar woodi
| Eschmeyer, W 1998: 1796 |
| Fowler, H 1951: 493 |
| Gosline, W 1945: 23 |
| Fowler, H 1941: 139 |
Hassar affinis
| Birindelli, J 2009: 276 |
| Ferraris Jr. C 2007: 171 |
| Eschmeyer, W 1998: 49 |
| Fowler, H 1951: 491 |
| Menezes, R 1949: 93 |
| Van der Stigchel, J 1946: 95 |
| Gosline, W 1945: 22 |
| Eigenmann, C 1925: 355 |
| Miranda Ribeiro, A 1911: 188 |
Hemidoras affinis
| Steindachner, F 1915: 73 |
| Eigenmann, C 1910: 394 |
| Eigenmann, C 1891: 33 |
| Eigenmann, C 1890: 258 |
| Eigenmann, C 1888: 158 |
Oxydoras affinis
| Steindachner, F 1881: 107 |