
Hassar gabiru, Birindelli & Fayal & Wosiacki, 2011
|
publication ID |
https://doi.org/ 10.1590/S1679-62252011000300006 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE1801-FFC4-FF82-D48B-F962DA89FB85 |
|
treatment provided by |
Carolina |
|
scientific name |
Hassar gabiru |
| status |
sp. nov. |
Hassar gabiru View in CoL , new species
Figs. 7 View Fig and 8 View Fig
Hassar orestis .— Camargo et al., 2004: 143 [in part, distribution, Xingu upstream, middle Xingu, Bacajá river].—Camargo, 2009: 208 [feeding habitat].—Camargo & Giarrizzo, 2009: 221 [biological data].
Holotype. MZUSP 108440 View Materials (1, 130.4 mm SL), rio Curuá , Iriri drainage, at Vila Castelo dos Sonhos, 8°19’7’’S 55°5’23’’W, Altamira, PA, Brazil, 22 Oct 2010, J. L. Birindelli, L. M. Sousa, A. L. Netto-Ferreira, M. H. Sabaj Pérez, N. Lujan. GoogleMaps
Paratypes. Xingu basin ( Brazil): ANSP 187335 About ANSP (3, 125.0- 151.7 mm SL), same data as holotype. INPA 26726 View Materials (1, 156.6 mm SL), rio Xingu , Ilha do Babaquara , Altamira, PA, Oct 1990, L. Rapp Py- Daniel & J. Zuanon. INPA 31043 View Materials (3, 122.5 mm SL [only one measured]), Mouth of rio Novo , 4°28’15’’S 53°40’6’W, Aug 2008, H. López-Fernández et al. MPEG 847 View Materials (3, 142.4- 172.2 mm SL), confluence of rio Culuene and rio Sete de Setembro , MT, Aug 1981, M. Goulding. MPEG 6736 View Materials (3, 108.7- 131 mm SL), Xingu basin, Altamira, PA, Jan 2001, Camargo. MZUSP 86888 View Materials (1, 56.2 mm SL), rio Sete de Setembro , 13°30’19’’S 52°24’57’’W, Canarana, MT, Oct 2004, C. R. Moreira et al. MZUSP 86991 View Materials (12, 66.5-91.4 mm SL), rio Culuene , 13°30’52”S 53°5’34”W, Gaúcha do Norte , MT, Oct 2004, J. L. Birindelli et al. MZUSP 87026 View Materials (2, 65.3-79.2 mm SL), rio Curisevo , Porto do Vitório , 13°2’5”S 53°25’19”W, Gaúcha do Norte , MT, Oct 2004, Moreira et al. MZUSP 94142 View Materials (6, 35.9- 136.0 mm SL), rio Culuene , 13°30’53’’S 53°5’40’’W, Gaúcha do Norte , MT, May 2007, J. L. Birindelli et al. MZUSP 94308 View Materials (1 c&s, 1, 114.0-116.0 mm SL), marginal lake, rio Culuene , 13°30’51’’S 53°5’49’’W, Gaúcha do Norte , MT, May 2007, J. L. Birindelli et al. MZUSP 94374 View Materials (4, 81.9-117.4 mm SL), lago da Mirim, rio Culuene, 13°25’48’’S 53°2’24’’W, Canarana, MT, May 2007, J. L. Birindelli et al. MZUSP 97219 View Materials (3, 124.7- 133.8 mm SL), same data as holotype. MZUSP 100328 View Materials (3, 60.4-65.9 mm SL), lagoas Tarik Yegun, rio Jatobá, 12°41’49’’S 54°9’8’’W, MT, Jul 2008, M. C. Lopes GoogleMaps .
Diagnosis. Hassar gabiru is distinguished from H. orestis and H. wilderi by having two weakly-branched diverticula restricted to each side of anterior chamber of gas bladder (rarely one extra pair on posterior chambers) (vs. gas bladder with many well-branched diverticula along margins of entire bladder); and gas bladder rounded posteriorly (vs. gas bladder triangular posteriorly, each posterior chamber extended posteriorly into a short terminal diverticulum sharing medial septum with its pair). Hassar gabiru is further distinguished from H. orestis by having 11 th through 15 th, modally 13 th, midlateral scute as the anteriormost with median thorn (vs. 1 st through 8 th, modally 3 rd), tip of upper caudal-fin lobe not darkened (vs. usually darkened), body depth at dorsal-fin origin 24.3-33.1%, mean 25.8% SL (vs. 16.8-22.2%, mean 21.1% SL), body depth at anal-fin origin 15.9-20.7%, mean 17.3% SL (vs. 10.0-14.7%, mean 13.1% SL), and caudal peduncle depth 6.6-8.6%, mean 7.1% SL (vs. 4.3-6.4%, mean 5.5% SL). Hassar gabiru is distinguished from H. affinis by having the distal tip of the first branched dorsal-fin rays and membranes pale (vs. first branched dorsal-fin rays and membranes distally darkened); and lateral diverticula on the gas bladder fingerlike, weakly-branched in large specimens (vs. rounded, rarely branched in large specimens).
Description. Morphometric data are summarized in Tables 1 and 2; holotype and additional specimens illustrated in Figs. 7 View Fig and 8 View Fig . Largest specimen examined 156.6 mm SL (INPA 26726). Dorsal profile of head rising moderately, evenly (usually in smaller specimens) or slightly convex (especially in larger specimens) from snout tip to anterior margin of orbit, and relatively straight form latter point to dorsal-fin spine. Dorsal profile of body descending gradually, approximately straight from dorsal-fin spine to caudal peduncle. Ventral contour shallowly concave from snout tip to pectoral girdle, and slightly convex from latter point to caudal peduncle. Caudal peduncle short with shallow hourglass shape in lateral view.
Body elongate with prominent conical snout. Mouth subterminal, each premaxilla bearing small patch of approximately 5 to 10 acicular teeth, and each dentary with approximately 10 to 20 acicular teeth. Oval orbit with weakly developed adipose eyelid in juveniles and adults, extended slightly beyond anterior and posterior limits of eye. Eyes positioned about half way between tip of snout and dorsalfin origin.
Three pairs of barbels (maxillary, inner and outer mental), all fimbriate. Maxillary barbel usually reaching base of first pectoral-fin ray; with 10 to 14 (mode 12, n=13) fimbriae along ventrolateral face. Inner and outer mentonian barbels of approximately equal size, covered with many rounded papillae, and falling short of ventralmost opening of gill slit.
Gill rakers small on first gill arch, absent on remaining arches. Accessory branchial lamellae on inner face of first gill arch well developed in approximately ten rows from insertion of rakers to origin of branchial filaments (but not contacting the latter); accessory lamellae gradually reduced on remaining (second to fifth) gill arches.
Lateral-line tubules ossified, forming a row of 30 to 33 (mode 31, n=13) midlateral scutes beginning with infranuchal. Three tympanal scutes, inconspicuous, usually without emergent thorn. Infranuchal scute with dorsal wing extremely thin and ventral wing dilated, expanded anteriorly, connected to posterior cleithral process; scute usually without medial thorn. Postinfranuchal scutes reduced anteriorly, nonoverlapping; each with posterior margin bicuspid (without medial thorn) or tricuspid (including medial thorn), latter condition usually starting at 13 th scute (range 11 th through 15 th, n=13); medial thorn and dorsal and ventral wings gradually increasing in size posteriorly; scutes with serrated posterior margins and overlapping on the posterior third of body.
Dorsal-fin II,6 (n=13), triangular with distal margin approximately straight, vertical when erected. Dorsal-fin spine slightly compressed and curved, with relatively small antrorse serrations along anterior margin (serrations reduced or absent on distal third); slightly larger retrorse serrations along posterior margin (serrations absent on proximal portion). Pectoral fin modally I,9, range I,8-9 (n=13); distal margin straight, oblique relative to body axis. Pectoral-fin spine slightly depressed and curved, with antrorse serrations along anterior margin (serrations absent on distal third); slightly larger retrorse serrations on posterior margin (serrations larger distally). Pelvic fin i,6 (n=13); distal margin rounded. Anal fin modally iv,9, range iii-iv,8-9 (n=13); subtriangular with scarcely rounded distal margin. Adipose fin relatively small, teardrop-shaped, with rounded free posterior margin. Caudal fin i,7/8,i (n=13, rarely i,7/7,i), distinctly forked, with lobes approximately equal in size.
Gas bladder ( Fig. 4 View Fig ) moderately large, cordiform. Gas bladder walls entirely smooth in small specimens (up to 50 mm SL). Large specimens have only two short weaklybranched diverticula on each side of anterior chamber; in the largest specimens examined a third pair of diverticula are present at middle of lateral margin of posterior chambers. Gas bladder rounded posteriorly, not extended into terminal diverticula.
Osteology. Osteology generally similar to that described for Hassar orestis , excepting differences as follows. Total vertebrae 35 (n=1), vertebrae 6-13 bearing ribs. Eight (n=1) dorsal-fin pterygiophores, 10 (n=1) pelvic-fin pterygiophores; 12 (n=2) dorsal and 12 (n=1) ventral caudal-fin procurrent rays. All examined specimens with hypural fusion pattern PH; HY 1+2; HY 3+4; HY 5, and with complete ossification of sutural joint at junction of sphenotic, parieto-supraoccipital and frontal.
Coloration. In alcohol, head and body tan to brown, or grey, countershaded. Faint dark stripe from anterior margin of upper lip to anterior margin of eye. A conspicuous dark blotch on the first three branched dorsal-fin rays and membranes, blotch starting from midlength of rays and membranes and almost reaching their distal tips, which are pale. Tip of upper caudalfin lobe not conspicuously darkened.
In life, ground color yellowish or greenish laterally and white ventrally; lower lip pinkish; eye silvery ( Fig. 8 View Fig ).
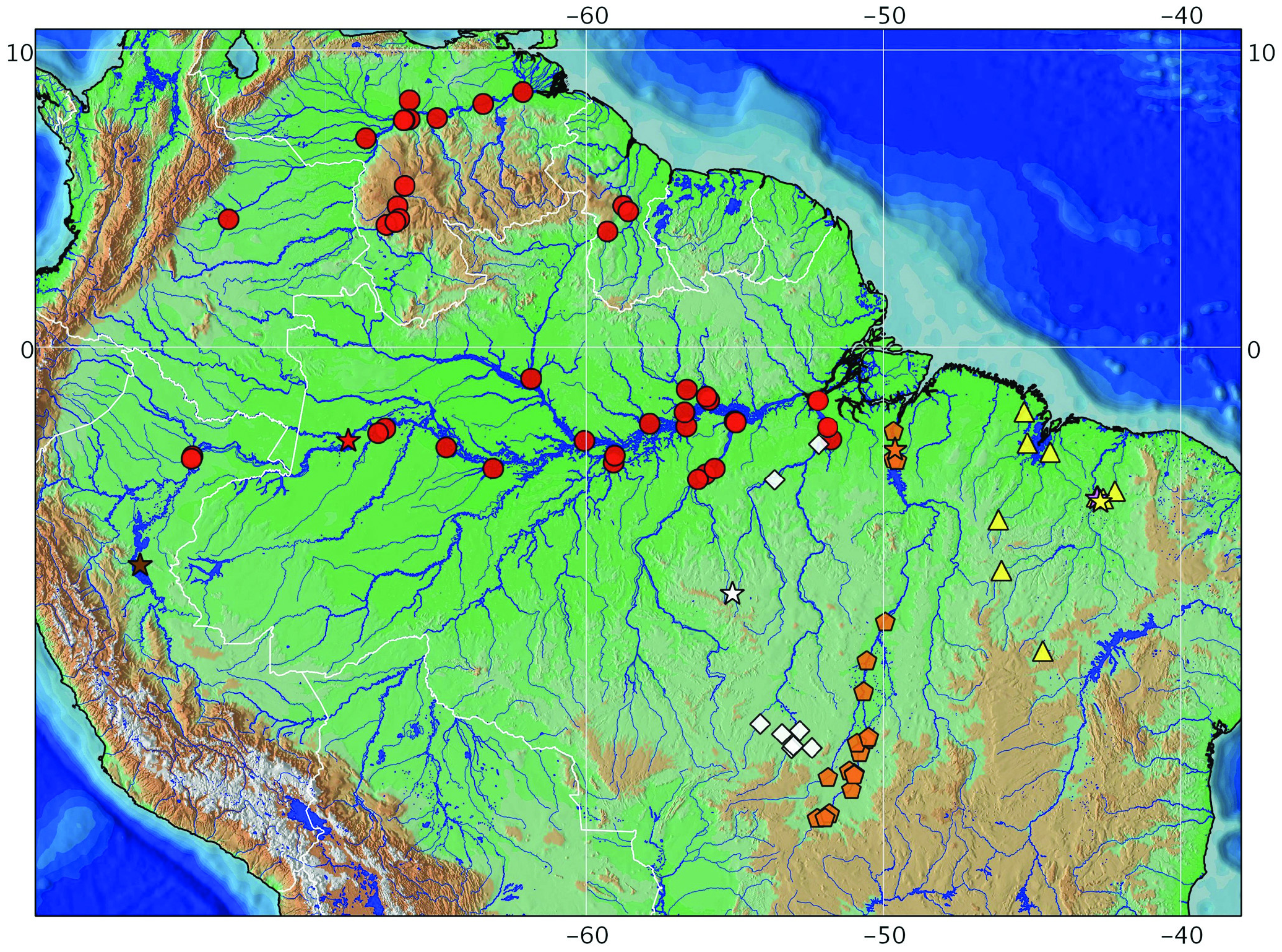
Distribution. Hassar gabiru occurs in the middle to upper Xingu river basin ( Fig. 6 View Fig ), and is apparently endemic to the Xingu basin above the rapids of Volta Grande, near Altamira.
Ecology. Like its congeners, Hassar gabiru was collected usually in swift water over sand beaches at night. Camargo (2009) considered that specimens of Hassar orestis (= Hassar gabiru ) collected at the Volta Grande rapids have preference for benthic invertebrates and detritus.
Etymology. Named in honor of Leandro Melo de Sousa, known to his friends as “Gabiru”, for his many contributions to the understanding of the Doradidae , including his MSc and PhD dissertations (Sousa & Rapp Py-Daniel, 2005; Sousa, 2010). Leandro also helped to collect part of the type series of the new species. Treated as a noun in apposition.
Remarks. Specimens of Hassar from the rio Xingu above the rapids of Volta Grande are distinct from the specimens below the rapids (including the area near Belo Monte) in overall body shape and gas bladder morphology, and are similar to the specimens from the upper Xingu river basin, herein referred to as the new species Hassar gabiru . This indicates that the Volta Grande rapids impose a distributional limit between H. gabiru and H. orestis . The identity of the specimens cited by Camargo et al. (2004), Camargo (2009) and Camargo & Giarrizzo (2009) were not verified by us. However, specimens collected at the rapids of Volta Grande or upstream (putatively including rio Bacajá) are likely to be Hassar gabiru .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |