
Maldybulakia Tesakov & Alekseev, 1998
|
publication ID |
https://doi.org/10.3853/j.0067-1975.50.1998.1288 |
|
DOI |
https://doi.org/10.5281/zenodo.4658230 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE7A7C-2D49-FFBB-FEAF-FA80FA467D73 |
|
treatment provided by |
Felipe |
|
scientific name |
Maldybulakia Tesakov & Alekseev, 1998 |
| status |
|
Maldybulakia Tesakov & Alekseev, 1998
1992 Lophodesmus Tesakov & Alekseev, 1992 View in CoL (non
Lophodesmus Pocock, 1894 View in CoL ).
Type species. Lophodesmus mirabilis Tesakov & Alekseev, 1992 .
Diagnosis. Large myriapodous arthropod with strongly mineralised pleurotergum, unmineralised sternum; cuticular surface with dense, polygonal sculpture; cuticle densely penetrated by large pore canals; trunk composed of two tagmata of presumed diplotergites; anterior tagma of one or possibly two subtrapezoidal tergite(s) having rounded corners; posterior tagma composed of at least four pleurotergites with pair of triangular lateral lobes on metazonites and short to long paratergal spines; pleurites coalesced with metazonites only; posterior-most (fifth) bilobate tergite with median spine-like process; spiracle in pleural furrow on third pleurotergite of posterior tagma; presumed telson composed of two small sclerites.
Discussion. Maldybulakia Tesakov & Alekseev, 1998 , was recently proposed as a replacement name for Lophodesmus Tesakov & Alekseev, 1992 , a name occupied by the extant polydesmid millipede Lophodesmus Pocock, 1894 . The generic diagnosis employed by Tesakov & Alekseev (1992) is modified to account for newly discovered diversity in the Devonian of Australia.
A major morphological contribution of new Australian species ofMaldybulakia is to elucidate the nature of trunk tagmosis in this arthropod. Tesakov & Alekseev (1992) identified two tergite types in beds containing Maldybulakia mirabilis . The type material consists of tergites regarded by them as diplosegments, with the lateral part of the "metazonite" swollen into a rounded lobe, and a spinose projection from the lateral edge of the paratergum. This tergite type will hereafter be referred to as a bilobate pleurotergite or B-pleurotergite (specimens of M. malcolmi n.sp. are interpreted as having pleurites fused to the tergum, thus the use of "pleurotergites").
Associated with the B-pleurotergites is a second, less common tergite type of "simple structure and roundedrectangular outline" ( Tesakov & Alekseev, 1992: 19). Noting that the rectangular (or, more accurately, trapezoidal) tergites bore a similar sculpture to the bilobate diplotergites/B-pleurotergites, Tesakov & Alekseev (1992) acknowledged the likelihood that they could belong to the same animal. However, they interpreted the trapezoidal tergites as neck segments, but (assuming one neck segment per animal, as for the dignathan collum) found them to be anomalously common relative to the typical B-pleurotergites.
Conclusive evidence that the trapezoidal tergites (here abbreviated T-tergites) are actually part of Maldybulakia is provided by the Australian species, in particular by several articulated specimens of M. malcolmi . Four articulated specimens of M. malcolmi have a single Ttergite attached or slightly displaced from the front of a series of B-pleurotergites. In the holotype, a T-tergite of nearly the same size as the articulated one (width 14.4 versus 14.8 mm, respectively) is displaced 14 mm from the front of the specimen. Considering the scarcity of specimens this small it is not unlikely that the displaced tergite belongs to the same individual, which would thus possess two T-tergites. The failure to find T-tergites in articulation with each other in any specimens may derive from a mainly membranous attachment and simple overlap (in contrast to the strong articulations developed between B-pleurotergites). Maldybulakia angusi reveals a range of morphology within this tergite type, suggestive of two or more T-tergites represented within a tagma of generally similar tergite form. The T-tergites vary in the presence or absence of a posteromedian spine, the prominence of tubercles, the degree of sinuosity of the transverse furrow, and the presence or absence of lateral swellings. Maldybulakia malcolmi and M. mirabilis display less morphological differentiation between the tergites of this tagma than is the case for M. angusi . This is consistent with evidence from the tagma composed of Bpleurotergites, wherein M. angusi shows a much greater degree of variation than seen in the other two species.
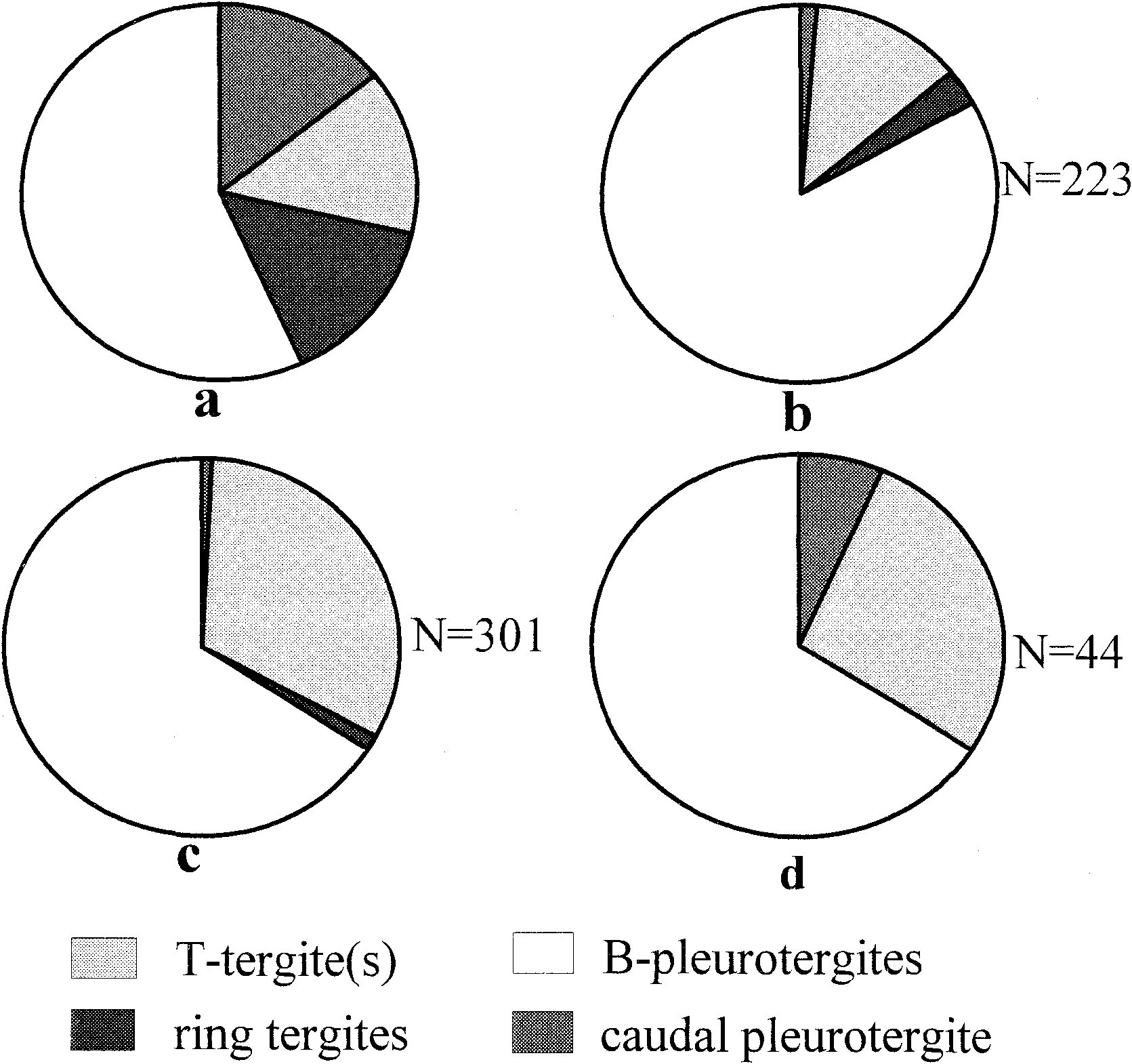
The frequency of occurrence of the trapezoidal tergites-27 percent of the sample (N=44)-was cited by Tesakov & Alekseev (1992) as a difficulty for their interpretation of the T-tergites as neck segments. Variable frequencies of occurrence of the T- and B-tergites are observed for the two Australian species of Maldybulakia ( Fig. 2 View Figure 2 ). The relative extent of the two tagmata might be estimated by the relative abundance of T- and B-tergites, although it appears that taphonomic factors have biased the samples. Three articulated specimens of M. malcolmi (AM F.102533, 102535, 102357) have a single T-tergite followed by four B-pleurotergites, then a caudal tergite as figured for M. mirabilis by Tesakov & Alekseev (1992). The caudal tergite is quite clearly a modified Bpleurotergite, showing the rounded lateral lobes and depressed, anterior overlapped surface (prozonite) typical of those pleurotergites. This serial homology is also obvious in M. malcolmi , in which the caudal tergite possesses small posterolateral projections as in the Bpleurotergites.
For Maldybulakia angusi , a survey of 301 sclerites that could be confidently identified.as either a T-tergite, ring tergite, B-pleurotergite or caudal tergite shows that Ttergites comprise 32 percent of the sample and Bpleurotergites 66 percent ( Fig. 2c View Figure 2 ). It might thus be inferred that the anterior tagmata composed of T-tergites is half the length of the tagmata comprising B-pleurotergites, which would accord well with the two T-tergites and four B-pleurotergites seen in the holotype of M. malcolmi . A ring-like tergite that overlies the prozonite of the anteriormost B-pleurotergite (and has processes permitting articulation of the T-tergite) in M. malcolmi is represented by only 4 of 301 specimens of M. angusi and is unreported in the sample of M. mirabilis ( Fig. 2d View Figure 2 ). Given that each individual possesses one of this sclerite type it is underrepresented in the sample. Caudal tergites are significantly underrepresented, known from only two specimens.
The relative abundance of disarticulated tergites in M. malcolmi differs from the 2:1 ratio of B:T tergites in the sample of M. angusi as well as the 4:1 or 4:2 ratio predicted by articulated specimens ( Fig. 2a,b View Figure 2 ). Of disarticulated tergites of M. malcolmi , 185 are B-p1eurotergites (83 percent) and 28 are T-tergites (12.6 percent). The apparent over-abundance of B-tergites is at the expense of ring tergites (3.1 percent) and caudal tergites (1.3 percent). These differences between the three species, if not entirely due to differential transport of the various sclerite types, might result from a greater number of T-tergites in M. angusi compared to M. malcolmi . There is a significant taphonomic bias observed in the case of articulation. Articulated specimens of M. malcolmi are smaller than most of the disarticulated material, presumably because larger specimens did not survive transport intact.
Although the trunk of Maldybulakia angusi displays much more serial variation than that of M. malcolmi , it has proven possible to identify most tergites of the former according to their position in the latter, and it cannot be ruled out that M. angusi may possess the same number of tergites in the trunk as known for M. malcolmi (one or possibly two T-tergites, a ring tergite, four B-pleurotergites and a caudal tergite). The minimal number of tergites in M. angusi is discussed more fully after description of that species.
Tesakov & Alekseev's (1992) evidence for regarding the B-pleurotergites of Maldybulakia mirabilis as diplosegmental (diplotergites) was their "distinct two-part structure". This refers to the separation of the anterior, articulating surface of the tergite from the posterior lobate part by a strong transverse groove or stricture. The Ttergites are of comparable proportions (length versus width) to the B-pleurotergites, and are also divided lengthwise by a transverse furrow in Australian species of Maldybulakia . This would suggest that they, too, would be diplotergites if the posterior trunk tergites were confirmed as being diplosegmental. All of the Palaeozoic myriapodous arthropods that might be compared with Maldybulakia on the basis of tergite form have proven to be diplosegmental when the appendages became known. Examples are kampecarids ( Almond, 1986), euphoberiid millipedes ( Burke, 1979), and arthropleurids ( Briggs & Almond, 1994). It is thus likely that the trunk tagmata of Maldybulakia are composed of diplosegments. The strong intertergal articulations and overlap of the prozonite of the B-pleurotergites by the preceding metazonite suggest that Maldybulakia had the capacity to enroll.
A pair of articulated sclerites are preserved just behind the caudal pleurotergite of the holotype of Maldybulakia malcolmi (Fig. 3a,g). They are also known from disarticulated specimens (Fig. 4g). Their positioning on the articulated specimen is a strong indication that they represent the posterior sclerites, although the nature of their (presumed) articulation to the caudal pleurotergite is not understood. Their small size, inferred posterior position, and overall structure invite a comparison with the telson of myriapods, which may also incorporate multiple sclerites (e.g., preanal sclerite or preanal ring, anal valves, and subanal plate in diplopods; Enghoff, 1990: 14-15). In the descriptions below, these two sclerites are called telson sclerites.
Information on pleural morphology is supplied by Maldybulakia malcolmi . The lateral exoskeletal component of Maldybulakia is regarded as a mineralised part of the pleuron (i.e., pleurites), rather than the tergum (paratergites) because ofits marked topographic separation from the tergum. Alternative interpretations of pleural structures are addressed under discussion of M. malcolmi .
The lack of sampling of a head in the Australian occurrences is curious, given the abundance of trunk tergites known for both species. Almond (1986) observed that articulated heads were rare in the possibly allied kampecarids, even when the trunk was fully articulated, and attributed this to a delicate attachment by arthrodial membrane, as is the case in millipedes. The head tergite of Kampecaris is aratherfeatureless plate (orpair of plates, the second possibly a collum according to Almond, 1986). The narrow sclerite alleged to be the head of Arthropleura ( Briggs & Almond, 1994: figs. 1, 2; Brauckmann et al., 1997) requires confirmation, as it may instead represent a collum-like tergite (W.A. Shear & H. Winkelmann, pers. comm., 1997). Even if the head of Maldybulakia likewise involved a simple plate it is unlikely that it has been overlooked because sclerites of this sort were deliberately sought, yet no candidates have appeared. Heads likely underwent a different transport history than the trunk, perhaps because of a membranous attachment. The possibility that specimens such as the holotype of M. malcolmi are complete, with the T-tergite being cephalic, is not favoured. This sclerite deviates from an expected morphology of an arthropod cephalon (e.g., lacking eyes or structures to accommodate them; lacking antennal sockets or notches), its transverse stricture and shape of the doublure conforming to a trunk diplotergite. The abrupt anterior termination of the doublure ( Fig. 6b View Figure 6 ) indicates a more anterior sclerite. The possibility that a species may possess two of these tergites further weakens the case for a cephalic identity. All of these factors outweigh the crude similarities in outline between a T-tergite and some arthropod head shields (e.g., the prosoma of bunodid xiphosurids, which has typical cephalic structures such as a cardiac lobe and ophthalmic ridges that are lacking in Maldybulakia ).
A few disarticulated sc1erites do not conform to those in articulated specimens of Maldybulakia malcolmi , and their position in the exoskeleton is unknown. A unique specimen of M. malcolmi (Fig. 5g,h) has a prominent, square embayment in could be presumed to be the posterior margin (this assuming that the strongly convex, ridge-like edge of the specimen is overlapped in articulation, as is the case for the prozonites). The conical median swelling warrants comparison with the caudal pleurotergite, although this swelling is strongly dorsally directed. The size and shape of the embayment invitespeculationthatthetelsonsclerites(Fig. 4g)might attach here. However, it does not seem possible that such a large, robust sc1erite could be positioned posteriorly in the trunk yet be missing from the holotype and other articulated specimens. As such, it is more likely situated anterior to the T-tergites, but attempts to interpret it as a head are unconvincing, lacking any landmarks indicative of an arthropod cephalic shield or head capsule.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Maldybulakia Tesakov & Alekseev, 1998
| Edgecombe, Gregory D. 1998 |
Lophodesmus
| Tesakov & Alekseev 1992 |
Lophodesmus
| Pocock 1894 |