
Torleya mikhaili, Tiunova, 1995
|
publication ID |
https://doi.org/10.11646/zootaxa.5418.1.6 |
|
DOI |
https://doi.org/10.5281/zenodo.10729749 |
|
persistent identifier |
https://treatment.plazi.org/id/03FFE945-FFB8-1F56-55CF-FC06D2E91BF4 |
|
treatment provided by |
Plazi |
|
scientific name |
Torleya mikhaili |
| status |
|
was first described by the author (1995) from the south of the Russian Far East, based on the male adult stage, reared from a larva ( Tiunova 1995). At that time, only four other species of the genus were known: Torleya nepalica ( Allen & Edmunds, 1963) from Nepal, T. major ( Klapalék, 1905) from Europe, T. padunica Kazlauskas, 1963 from eastern Siberia and T. japonica Gose, 1980 from Japan. In 2004, a review of Torleya Lestage species was carried out that included eight valid Torleya species from the Palaearctic and Oriental Regions (Jacobus et al. 2004). The previous five species and three others were included: Torleya grandipennis Zhou, Su & Gui, 2000 , T. lutosa Kang &Yang, 1995 , and T. naga Jacobus &McCafferty (in Jacobus et al.), 2004. Later, in a work devoted to the revision of Ephemerellidae genera, three more species were added. Torlyea coheri ( Allen & Edmunds, 1963) and T. lacuna ( Jacobus, McCafferty & Sites, 2007) were moved from the genus Crinitella Allen & Edmunds , and T. longforceps ( Gui, Zhou & Su, 1999) was transferred from the genus Serratella Edmunds. Torleya longforceps was pleased in this genus on the basis of its male genitalia being similar to those of T. coheri , T. lacuna , and T. mikhaili . At the same time, T. grandipennis was moved to the genus Hyrtanella Allen & Edmunds ( Jacobus & McCafferty 2008) . Currently, the genus Torleya includes 13 species. This includes two species based on larvae from India— T. simbalbarensis Selvakumar, Subramanian, Martynov & Jacobus (in Martynov et al.), 2021 and T. dibruensis Selvakumar, Martynov & Jacobus (Martynov et al.) 2021 —and T. elissa ( Jacobus, Zhou & McCafferty 2009) from Iran, which was moved from Serratella by Martynov et al. (2023).
The increased number of species in the genus Torleya over the last thirty years has been associated with the description of new species and the movement of some species from other genera. Thus, a number of morphological characters that are necessary for a diagnosis of species are often absent from earlier descriptions (Jacobus et al. 2004). Considering that since the description of T. mikhaili the author has collected new material of both larvae and adults from different regions of the Russian Far East, it was decided to present additional characteristics and original illustrations for larvae and adults, as well as to describe for the first time the female imago and eggs.
Material examined. Russian Federation: Primorye Territory: Holotype. ♂ imago (reared from larva). Ussuri River, 1.5 km below Utyos Mountain, 02.VIII.1992, T. Tiunova.
Paratypes. 5♂ 1♀ imagoes, 5♂ 1♀ subimagoes, all reared from larvae, 4 larvae, Ussuri River , 1 km below Utyos Mountain, 04.VIII.1994 ; 5♂ imagoes, 2♂ subimagoes, all reared from larvae, 1 larva, Ussuri River, Gornye Kluchi Village , 2 km below bridge, highway Vladivostok-Khabarovsk 18.VII.1996 ; 1♂ imago,1 larval skin ♀, 1 larval skin ♂, Bolshaya Ussurka River , 1 km below Zvenigorogka Village, 20.VII.1996 ; 13 larvae, Bolshaya Ussurka River basin, Malinovka River , 50 m above bridge, highway Vladivostok-Khabarovsk, 20.VII.1996 ; 8 ♂ imagoes (light), Ilistaya River , 700 m below bridge, highway Vladivostok-Khabarovsk, 5.VIII.2007 ; 1♂ imago (light), Razdol’naya River, above Zarechnoe Village , 4.VIII.2007 ; 2♂ imagoes (light), Razdol’naya River Basin, Medveditsa River at the confluence with the Kraunovka River , 14–15.VIII.2002 ; Khabarovsk Territory: 1♂ (reared from larva), Amur River Basin, Bikin River, Zvenevoy Village , 22.VII.1996 ; 13 larvae, Bikin River , 300 m below bridge, highway Vladivostok-Khabarovsk, 13.VII.2000 ; 1♂ (reared from larva), Amur River Basin, Gur River , bridge, highway Khabarovsk-Komsomolsk, 22.VII.1996 . All material collected by T. Tiunova.
Diagnosis. Torleya mikhaili can be separated from the other species of the genus by the combination of the following characteristics. Male imago: (1) forceps segment II which has a slight distal expansion; (2) penis with elongated apical lobes rounded at the apex; (3) U-shape a median notch; (4) genital forceps segment III relatively short, length approximately 1.5–1.7 times the width. Larvae: (5) posterior margins of abdominal terga without paired tubercles; (6) forefemur with distal transverse row of long, stout and pointed setae; length of these setae ½ of width of femur; (7) claw of fore leg with two groups of denticles; apical group with four acute and long denticles; subdistal setae absent; (8) posterolateral projection on terga IV–IX well developed; (9) posterior margins of terga IV–IX with paired submedian short, clavate setae. Eggs: (10) the ridges about 2–3 µm in width; (11) the surface of the ridges relatively smooth.
Redescription
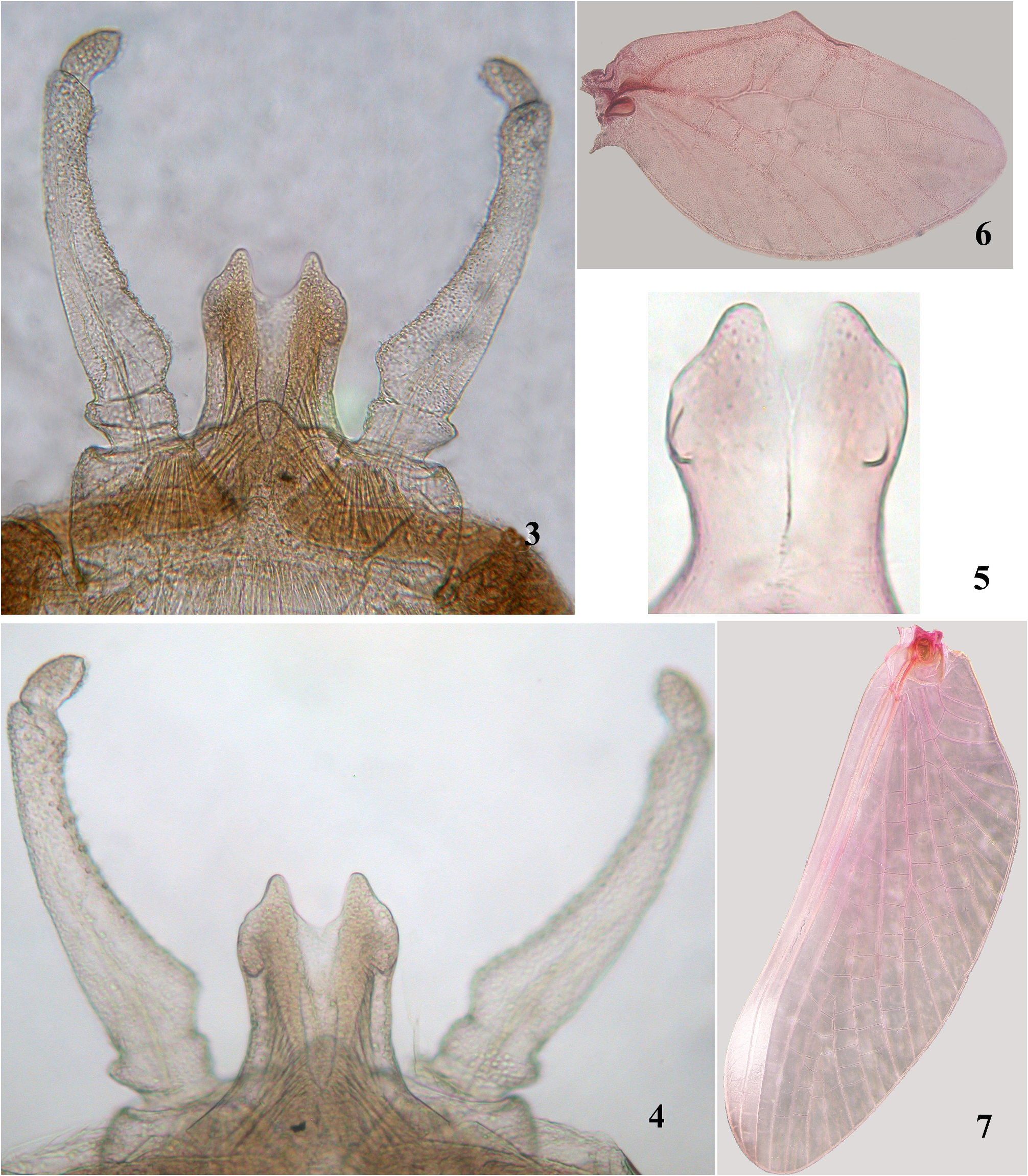
Male imago (in alcohol) ( Fig. 1 View FIGURES 1–2 ). Length (mm): body, 4.7–5.6; fore wings, 4.6–5.6; caudal filaments, 4.5–5.3. Length of segments in foreleg (mm): femora 0.95–1.0, tibia 2.3–2.6, tarsal segments 0.45–0.5, 0.35–0.4, 0.2–0.25, 0.15–0.2, 0.1. Abdominal terga I–IV or I–VII dirty brownish, translucent; tergum V–IX or VII–IX light brown, opaque ( Fig. 1 View FIGURES 1–2 ). Wings hyaline with translucent milky tinge along costal margin; longitudinal veins, intercalaries, and crossveins hyaline ( Figs 6–7 View FIGURES 3–7 ). Second segment of genital forceps with slight distal expansion ( Figs 3–4 View FIGURES 3–7 ); third segment length approximately 1.7x width. Penis lobes slightly elongated and rounded at apex and with U-shaped, shallow and relatively wide median notch ( Figs 3–4 View FIGURES 3–7 ); penis lobes with small subapical projection ( Fig. 4–5 View FIGURES 3–7 ). Caudal filaments pale ( Fig. 1 View FIGURES 1–2 ).
Female imago (in alcohol, reared from larva) ( Fig. 2 View FIGURES 1–2 ). Length (mm): body 4.7; fore wings, 5.0; caudal filaments, 4.5. Head, thorax and abdomen yellowish brown; thorax somewhat darker than abdomen. Wings hyaline, all veins pale. Legs pale. Abdomen with eggs yellowish or brownish; abdominal sterna yellowish. Subanal plate triangular with rounded apex. Caudal filaments pale.
Male subimago (in alcohol) ( Fig. 8 View FIGURES 8–9 ). Similar to male imago, but color of abdomen uniformly yellowish-brownish.
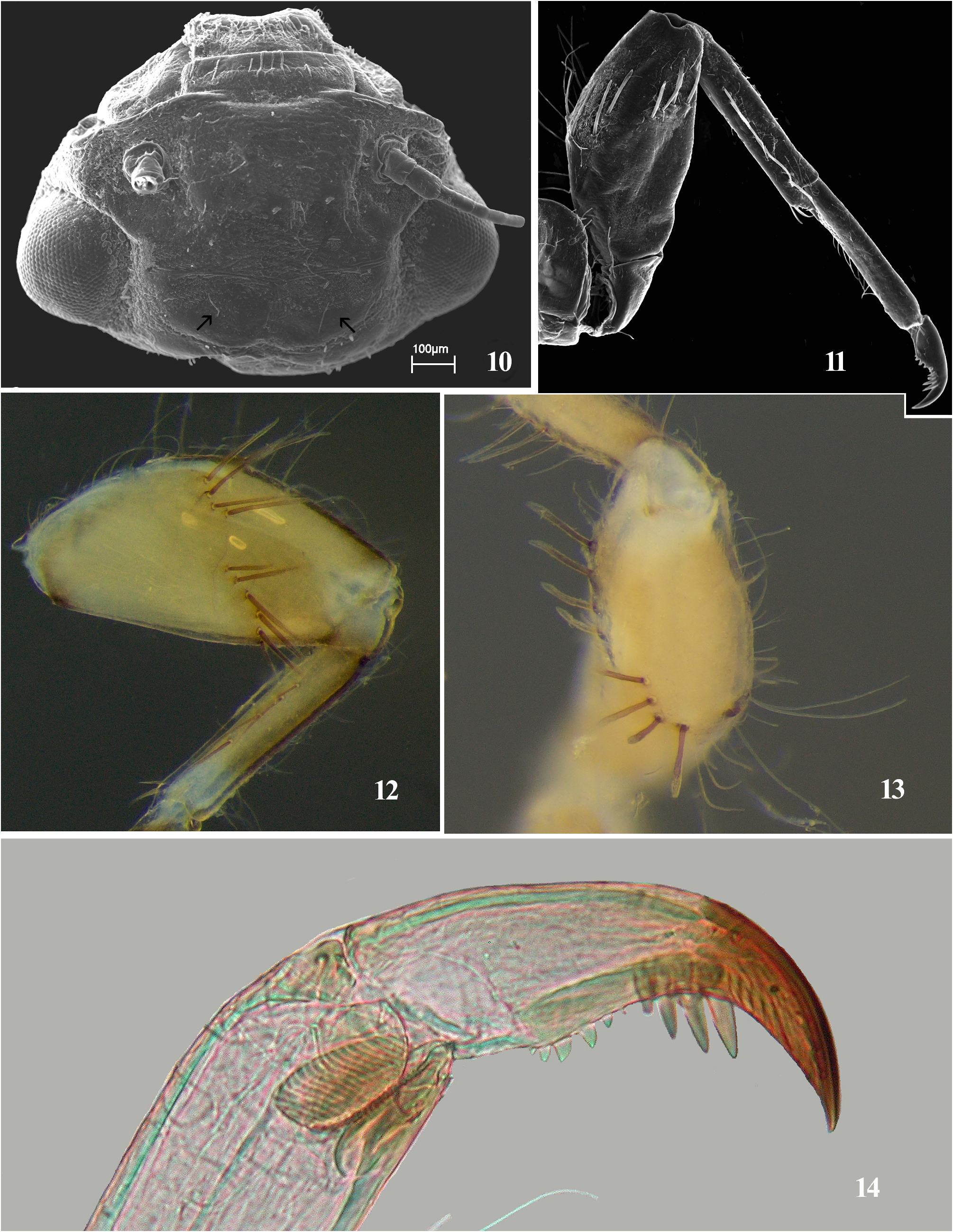
Final larval instar (in alcohol) ( Fig. 9 View FIGURES 8–9 ). Length (mm): body, 3.8–5.5; caudal filaments: 1.4–2.3. General colouration light brown with brown markings ( Fig. 9 View FIGURES 8–9 ). Head with pair of long hairlike setae on occiput ( Fig. 10 View FIGURES 10–14 ). Mouthparts (see also Tiunova 1955: figs 3–5): clypeus with anterior row of long, stout, hairlike setae ( Fig. 10 View FIGURES 10–14 ); dorsal surface of labrum densely covered with long, fine setae ( Fig. 10 View FIGURES 10–14 ). Maxilla without palp (see Tiunova 1955: fig. 4). Pronotum without any protuberances ( Fig. 9 View FIGURES 8–9 ). Forefemur with distal transverse row of long, stout and pointed setae ( Figs 11–13 View FIGURES 10–14 ); length of these setae ½ of width of femur ( Figs 11–12 View FIGURES 10–14 ); mid and hind femora each with longitudinal, sparse row of long thick setae and long thin hairlike setae along dorsal and ventral margins ( Fig. 9 View FIGURES 8–9 ). Fore tibia with row of stout, brown setae and pale hairlike setae along ventral margin; mid and hind tibia and tarsi with long, thin hairs along dorsal and ventral margins ( Fig. 9 View FIGURES 8–9 ). Claw with two groups of denticles; apical group with four acute and long denticles, gradually longer apically; basal group with four or five blunt denticles, middle denticles largest; subdistal setae absent ( Fig. 14 View FIGURES 10–14 ). Lengths (mm) of leg segments. Fore leg: femur 0.7–0.75; tibia 0.5–0.55; and tarsus 0.45–0.5. Middle leg: femur 0.75–0.85; tibia 0.5–0.6; and tarsus 0.45–0.5. Hind leg: femur 0.85–0.9; tibia 0.6–0.75; and tarsus 0.5–0.6. Posterior margins of terga IV–IX with paired submedian short, clavate setae, which may be single on terga IV and IX, tuft of two or three setae on terga V–VII, and row of three or four setae on tergum VIII which continues to lateral margin ( Fig. 15 View FIGURES 15–18 ); terga II–III without posterolateral projections; posterolateral projections on terga IV–IX well developed ( Fig. 15 View FIGURES 15–18 ); anterolateral margin of tergum II with tuft of long, hairlike setae; lateral margins of terga IV–IX each with row of long, stout setae of various sizes; lateral setae on tergum IX shorter than those on other terga ( Fig. 9 View FIGURES 8–9 ). Caudal filaments about one fourth as long as body; setae at apex of each segment; setae of different shapes: long and hairlike, short stout, oval ( Figs 17–18 View FIGURES 15–18 ). Distal margins of lamellate portion of gills III–V with shallow notch in middle part and rounded protrusion on outer side ( Figs 19–22 View FIGURES 19–22 ); gill III lamella largest, slightly tapering distally without any weak transverse band ( Fig. 19 View FIGURES 19–22 ); lamellae of gills IV and V almost same shape, with strongly attenuated, rounded anterior outer margin ( Figs 20–21 View FIGURES 19–22 ); lamella of gill VI almost symmetrical in shape, with rounded distal margin, and well-defined rounded projection on anterior edge of outer margin ( Fig. 22 View FIGURES 19–22 ); gill VII transparent, tiny, with weakly defined attachment.
Eggs. General form of egg ovoid with length of 129–145 µm and width of 75–85 µm ( Figs 23–24 View FIGURES 23–26 ); polar cap present, composed of dense threads ( Fig. 26 View FIGURES 23–26 ). Chorion with geometric macrorelief constituted of large-meshed reticulations with relatively deeply mesh units; each mesh unit with flat, smooth bottom; ridges about 2–3 µm in width; surface of ridges relatively smooth; inner margin of ridges with numerous papillae ( Fig. 25 View FIGURES 23–26 ). Micropyles in subequatorial area; sperm guide rounded or teardrop-shaped ( Fig. 25 View FIGURES 23–26 ).
Habitat. Larvae and adults of T. mikhaili were collected in the south of the Russian Far East (Primorskiy and Khabarovskiy Territories) at the beginning July–middle of August. Larvae were collected in rivers ( Figs 27–28 View FIGURES 27–28 ) at a depth of 30 to 100 cm. The width of the rivers in the places where larvae were collected exceeded 100 m; the length of these rivers is more than 300 km. The water temperature in this period exceeds 20–25°C. The larvae inhabited stone-gravel substrates with sand and detritus, mainly in slow currents.
Distribution. East Palaearctic: the south portion of the Russian Far East, northeastern China (Jacobus et al. 2004; Li et al. 2020).
Remarks. In an article devoted to the revision of Torleya species, the authors were the first to indicate China in the distribution of T. mikhaili (Jacobus et al., 2004) . At the same time, when comparing larvae from Russia and China, a number of differences were noted that arose due to the absence of some morphological characteristics in the description of T. mikhaili . Additional data presented in this work confirm the presence of this species in China. Note that in 2020 an article was published that provided molecular data for T. mikhaili larvae collected in China ( Li et al. 2020). Therefore, a goal of further research will be to collect material for the molecular study of T. mikhaili from Russia so those populations can be compared to their Chinese counterparts.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |