
Pristimantis yantzaza, Valencia & Dueñas & Székely & Batallas & Pulluquitín & Ron, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4353.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:A4FD63BD-47ED-4B4B-BC65-5E75738DDC89 |
|
DOI |
https://doi.org/10.5281/zenodo.6000667 |
|
persistent identifier |
https://treatment.plazi.org/id/04058030-4A17-206D-FF66-F8C7B930F9F1 |
|
treatment provided by |
Plazi |
|
scientific name |
Pristimantis yantzaza |
| status |
sp. nov. |
Pristimantis yantzaza sp. nov.
Suggested English name: Yantzaza´s Rainfrog Suggested Spanish name: Cutín de Yantzaza ( Figs. 2A–B View FIGURE 2 )
Holotype. FHGO 9962 ( Figs. 2–3 View FIGURE 2 View FIGURE 3 ) an adult female from La Zarza , Los Encuentros, Yantzaza, (3°46ʼ36” S, 78°29ʼ47” W, 1478 m elevation), Zamora Chinchipe province, Republic of Ecuador, collected on 12 October 2014 by M. R. Dueñas and D. Hurtado.
Paratopotypes ( n =6). DHMECN, 11696, 11697, an adult male and an adult female collected on 12 December 2012 by M. R. Dueñas; FHGO 9963–65 an adult male and two adult females with the same collection data as the holotype. FHGO 8346, an adult female, collected on 25 February 2011 by A. Barahona and H. Cadena.
Paratypes ( n = 26). DHMECN, 11693 an adult female and DHMECN 11700, 11702 two juvenile males from Colibrí, Los Encuentros (3°45ʼ47” S, 78°30ʼ13” W, 1680 m), Zamora Chinchipe province, collected on 12 December 2015 by M. R. Dueñas; FHGO 8346, 9963, two adult females from Colibrí, Relaves, Yantzaza (3°45ʼ42” S, 78°30ʼ16” W, 1390 m), Zamora Chinchipe province collected on 25 February 2011 by A. Barahona and H. Cadena. DHMECN, 8504, an adult female, ( 3°34'31” S, 78°26'10” W, 1346 m), collected on 2 December; DHMECN 10157, an adult female, ( 3°31'55” S, 78°23'43” W, 926 m), collected on 6 December 2011; DHMECN 11147, an adult female ( 3°34'28” S, 78°26'04” W, 1253 m), collected on 23 June 2013; DHMECN 11149, 11153– 54, 11159, three adult males and an adult female (3°35ʼ07” S, 78°26ʼ22” W, 1590 m), collected on June 2013, all from Concesión Mirador, Tundayme, Pangui, Zamora Chinchipe province collected by R. M. Betancourt, D. Chungandro and M. Alcoser; FHGO 8884, 8886, two adult female from Las Peñas, Relaves, Yantzaza ( 3°46'25” S, 78°29'43” W, 1480 m), Zamora Chinchipe province, collected on 4 November 2012 by J. H. Valencia; QCAZ 63294, 63295, two adult females from 20 km E Santa Clara del Vergel, Palanda (4°36ʼ41” S, 78°54ʼ12” W, 2030 m), Zamora Chinchipe Province, Ecuador, collected on 24 March 2016 by P. Székely. Ten adult males, QCAZ 65056 (4°35ʼ54” S, 78°51ʼ34” W, 1644 m), QCAZ 65063 (4°36ʼ14” S, 78°49ʼ42” W, 2112 m), QCAZ 65096, 65098–99, 65102, (4°36ʼ6” S, 78°49ʼ51” W, 2033 m), QCAZ 65110 (4°36ʼ1” S, 78°49ʼ57” W, 2014 m), QCAZ 65112 (4°36ʼ2” W, 78°49ʼ56” W, 2016 m), QCAZ 65143 (4°35ʼ57” S, 78°50ʼ4” W, 1981 m), QCAZ 65147 (4°35ʼ40” S, 78°50ʼ25” W, 1832 m), collected on 20–27 August 2016, all from Reserva Biológica Cerro Plateado, La Canela, Palanda, Zamora Chinchipe province, by D. Almeida, E. Nursirquia, F. Ayala, J. Pinto, A. Achig, and M. Bustos.
Referred material ( n = 12). DHMECN, 11693 (3°45ʼ03” S, 78°30ʼ40” W, 1620 m) an adult female and DHMECN 11700 (3°45ʼ47” S, 78°30ʼ13” W, 1680 m) a juvenile male, both collected on 12 December 2012 from Colibrí, Los Encuentros, Zamora Chinchipe province by M. R. Dueñas. DHMECN 11150, 11159, two adult females (3°34ʼ33” S, 78°26ʼ43” W, 1250 m), DHMECN 11157, 11160 an adult male and an adult female (3°34ʼ47” S, 78°26ʼ22” W, 1361 m), all collected on June 2013 from Concesión Mirador, Tundayme, Pangui, Zamora Chinchipe province collected by R. M. Betancourt and D. Chungandro. Seven adult males QCAZ 65057 (4°35ʼ40” S, 78°51ʼ14” W, 1619 m), QCAZ 65095 (4°36ʼ6” S, 78°49ʼ51” W, 2033 m), QCAZ 65103 (4°36ʼ11” S, 78°49ʼ40” W, 1996 m), QCAZ 65119 (4°36ʼ10” S, 78°49ʼ48” W, 2073 m), QCAZ 65146 (4°35ʼ57” S, 78°50ʼ6” W, 1980 m), QCAZ 65148, 65150 (4°35ʼ38” W, 78°50ʼ31 W, 1803 m), collected on 20–27 August 2016, all from Reserva Biológica Cerro Plateado, La Canela, Palanda, Zamora Chinchipe province, by D. Almeida, E. Nursirquia, F. Ayala, J. Pinto, A. Achig, and M. Bustos.
Comparisons with other species. Coloration in this section refers to live specimens unless otherwise noted. Pristimantis yantzaza sp. nov. is readily distinguished from other congeneric species in Cordillera del Condor by its bright reddish, salmon or orange ventral coloration and iris cream bluish to light blue with dark reticulations and a wide dark red mid-horizontal band. Ventral coloration is shared with P. paquishae (Brito et al. 2014) and Pristimantis proserpens ( Lynch 1979) , however P. paquishae has vocals slits and short and robust digits, while P. proserpens lacks discoidal folds and has longer digits. In Cordillera del Condor there are fifteen other species of Pristimantis (Almendariz et al. 2014) that could be related and or confused with P. yantzaza (characters of the new species given in parenthesis). Pristimantis croceoinguinis ( Lynch 1968) and P. ventrimarmoratus ( Boulenger 1912) lack tympanic annulus and tympanic membranes (annulus and membrane tympanic present); P. trachyblepharis ( Boulenger 1918) and P. spinosus ( Lynch 1979) have tympanum concealed beneath skin, length 30–40% eye length (annulus and membrane tympanic distinct, 40% eye length), P. prolatus ( Lynch & Duellman 1980) , P. proserpens ( Lynch 1979) , and P. versicolor ( Lynch 1979) lack discoidal fold (discoidal fold present); Pristimantis altamazonicus ( Barbour & Dunn 1921) , P. serendipitus ( Duellman & Pramuk 1999) , and P. exoristus ( Duellman & Pramuk 1999) have vocal slits in males (vocal slits absent); P. diadematus (Jimenez de la Espada 1875) has a tarsal fold (tarsal folds absent); P. muscosus ( Duellman & Pramuk 1999) has reddish iris (iris cream bluish to light blue); Pristimantis minimus Terán – Valdez & Guayasamin 2010, is a minute frog, SVL 12.2 mm in males and 9.5– 13.7 mm in females (SVL 17.6–22.0 in males, 21.6–34.0 in females); P. rhodostichus ( Duellman & Pramuk 1999) , has fingers bearing distinct lateral fringes (fingers bearing narrow lateral fringes), has low cranial crests (cranial crests absent). Pristimantis tungurahua Reyes –Püig et al. 2010, distributed on eastern Andean slopes, also have reddish venter coloration; however P. tungurahua has dorsolateral folds, subacuminate snout in dorsal view and acuminate in lateral view, and heel with prominent calcar. Additionally, P. yantzaza can be easily distinguished from all the other phylogenetically close species, P. ardalonychus ( Duellman & Pramuk 1999) , P. cajamarcensis ( Barbour & Noble 1920) , P. ceuthospilus ( Duellman & Wild 1993) , P. minutulus Duellman & Hedges 2007 , P. ockendeni ( Boulenger 1912) , and P. unistrigatus ( Günther 1859) , by its characteristic reddish, salmon or orange ventral coloration.
Etymology. The specific epithet yantzaza refers to canton of Yantzaza, one of the eight cantons of the province of Zamora Chinchipe, where the new species is known to occur. In addition, Yantzaza is an Ecuadorian Shuar word that means “firefly”. Yantzaza was traditionally called “Firefly Valley” or “Valle de Yantzaza” by indigenous Shuar people. The epithet is a noun in apposition.
Diagnosis. We assign this species to the genus Pristimantis on the basis of its phylogenetic position. The new species also shows a general morphological similarity with many other members of the genus (including characteristic T–shaped terminal phalanges, toes without membranes, and Toe V longer than Toe III). Pristimantis yantzaza sp. nov. is characterized by: (1) skin of dorsum finely shagreen with scattered pustular tubercles, dorsal folds absent; skin of belly coarsely areolate, discoidal folds weak; thoracic fold present; (2) tympanic membrane and tympanic annulus evident, round; horizontal diameter of tympanic annulus 45–50% of eye diameter and separated from the eye by a distance of approximately one third of the length of the eye; (3) snout short, subacuminate in dorsal view, rounded in profile, canthus rostralis distinct, rounded in dorsal view and slightly sharp in lateral profile, loreal region slightly concave to flat; nostrils slightly protuberant, directed dorsolaterally; area between nostril slightly convex; (4) upper eyelid bearing several small and flat tubercles and two or three enlarged and sub–conical supraocular tubercles, approximately 60% of eye diameter; cranial crests absent; (5) choanae triangular not concealed by palatal shelf of maxillary arch; dentigerous processes of vomers small and ovoid; slightly smaller than choanae, separated medially by distance lower than width of odontophore; each odontophore has 5 to 7 teeth; (6) males lacking vocal sac and slits; (7) Finger I shorter than Finger II, all fingers bearing subtruncate disks, approximately three times the width of the digit proximal to the pad; (8) fingers bearing narrow lateral fringes; (9) small ulnar tubercles present; (10) inner edge of tarsus bearing indistinct tubercles, heel bearing one large tubercle and several smaller tubercles; tarsal fold absent; (11) inner metatarsal tubercle ovoid and slightly elevated, about four times the diameter of the outer tubercle; outer metatarsal tubercle small, sub-conical; supernumerary plantar tubercles present; (12) toes bearing narrow lateral fringes; Toe IV much longer than Toe III; discs broadly expanded, elliptical, about same size as those on fingers; (13) dorsum yellowish tan to dark brown, mottled with darker brown or with cream vertebral and dorsolateral stripes; concealed surface of thigh reddish, salmon or orange with brown bars; venter and groin uniformly bright reddish or orange, throat bearing minutes spots; (14) SVL in adult males 17.6–22.1 mm (n = 6), females 23.2–34.0 mm (n = 16).
Description of holotype. Adult female ( Figs. 3A–D View FIGURE 3 ) with, 24.4 mm SVL; head slightly wider than body; head barely wider than long; head length 41.9% of SVL. Canthus rostralis sharp, loreal region slightly concave to flat, sloping abruptly; nostrils slightly protuberant, directed laterally; eye-to-nostril distance 47.7% of eye diameter; lips not flared; upper eyelid bearing several small and flat tubercles and two enlarged and sub-conical supraocular tubercles; snout rounded in dorsal and lateral view; tympanic membrane and tympanic annulus evident, round, its upper and posterodorsal part obscured by rounded supratympanic fold; supratympanic fold thick and slightly warty; several low postrictal tubercles situated posteroventrally to tympanic annulus; interorbital distance greater than the width of upper eyelid; choanae small, rounded, not concealed by palatal shelf of maxillary arch; dentigerous processes of vomer small and oblique with four teeth; tongue longer than wide, bilobate, posterior 1/3 not adherent to floor of mouth.
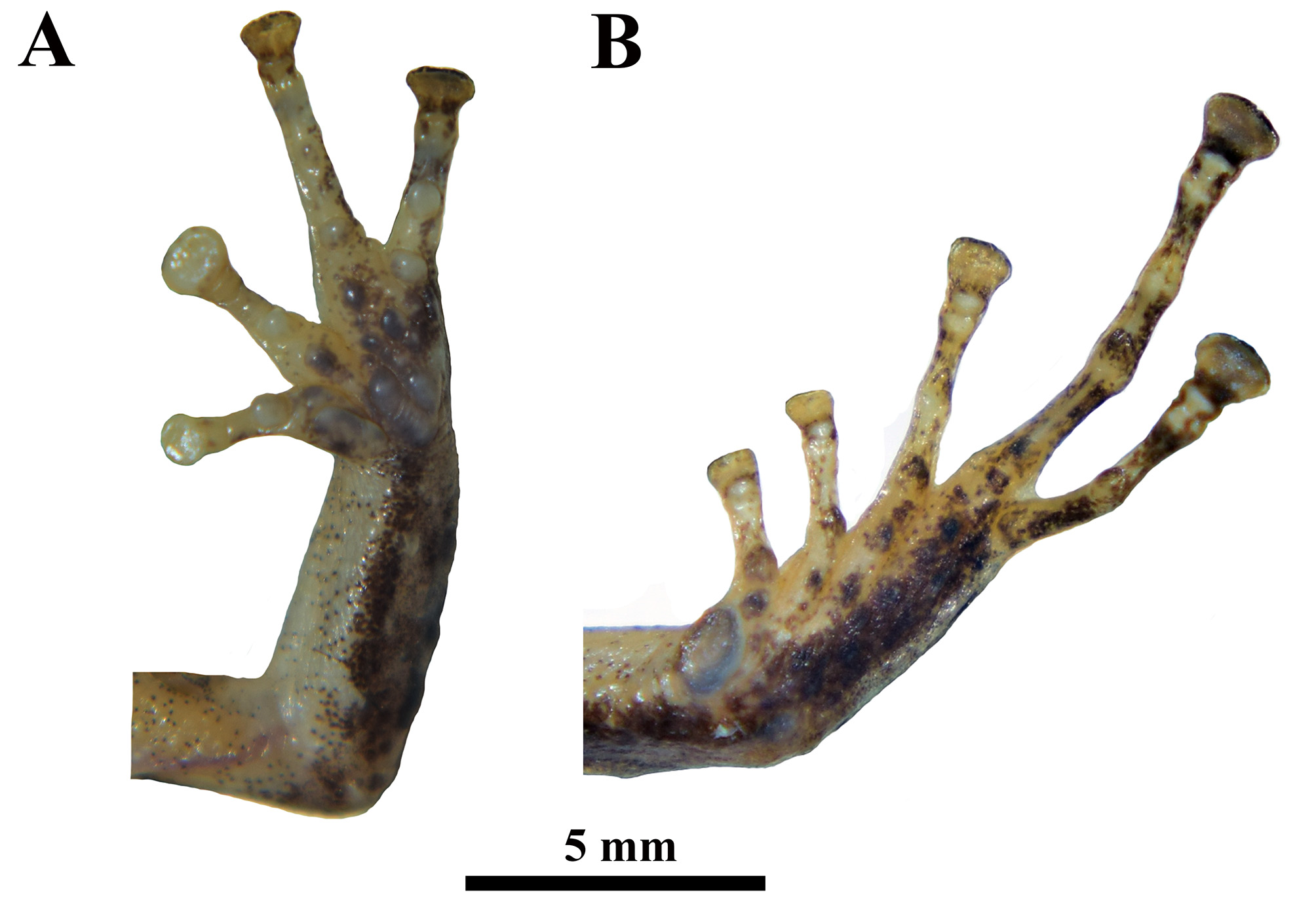
Skin of dorsum finely shagreen; dorsolateral folds absent; skin of throat shagreen; thin, low mid dorsal fold starting at tip of snout and ending at cloaca (trait more visible in life); low interocular fold present (trait more visible in life); ventral and lateral skin coarsely areolate; discoidal folds weak; thoracic fold present (trait more visible in life); small ulnar tubercles present without forming a distinct fold; thenar tubercle oval equal (right hand) or barely smaller (left hand) than divided palmar tubercle; supernumerary palmar tubercles present; subarticular tubercles prominent and rounded, including the most distal tubercle; fingers bearing narrow lateral fringes, all fingers bearing pads well defined by circumferential grooves; Finger I shorter than Finger II; tips of digits subtruncated; inner digit of hand bearing barely narrower disc than outer digits ( Fig. 4A View FIGURE 4 ); all fingers bearing pads on digital tips; tips of Finger IV reaches beyond distal subarticular tubercle on Finger III; tip of Finger II reaches distal subarticular tubercle of Finger III; knee bearing low tubercles.
Hind limbs moderately long; tibia long 56.5% of SVL; foot long 46.1% of SVL; Toe V much longer than Toe III; tip of Toe III not reaching the distal edge of the penultimate subarticular tubercle on Toe IV; tip of Toe V extending beyond the distal edge of distal subarticular tubercle on Toe IV ( Fig. 4B View FIGURE 4 ); disks of inner toes slightly smaller than outer fingers; tips of digits subtruncated; webbing absent; subarticular tubercles on toes distinct, projecting and ovoid; supernumerary plantar tubercles at base of toes present; inner metatarsal tubercle ovoid and elevated, almost half the length of Toe I; lateral fringes absent; outer metatarsal tubercle small, sub–conical; tarsus without fold but small sub–conical tubercles along outer border, indistinct on inner border; heel bearing one larger, rounded tubercle and several smaller tubercles; cloacal sheath and tubercles present, cloacal opening directed posteroventrally at level of thighs.
Coloration in life. Dorsum is yellowish–orange with brown-reddish markings; suprascapular region bearing W markings; dark brown interorbital bar; brown cloacal region. Dorsal surfaces of limbs yellowish-orange with dark brown bars. Dorsum of hands and feet with brown blotches, more abundant on outer fingers and toes; fingers I and II pale reddish–brown. Flanks bright yellowish with reddish-brown markings. Belly, axial region and groin bright reddish lacking spots or blotches, reddish throat finely mottled with minute brown spots. The concealed surfaces of limbs are reddish-brown finely mottled with pale brown. Ventral surfaces of tarsus dark brown. Palmar surface reddish densely mottled with brown, subarticular, palmar and thenar tubercles reddish; dark brown bars originating below the eyes and extending towards the lips. Iris bright creamy with a reddish median, horizontal streak and faint black reticulations ( Figs. 2A–B View FIGURE 2 ).
Coloration in preservative. Dorsum is pale brown with dark brown markings; dark brown interorbital bar; brown cloacal region. Dorsal surfaces of limbs pale brown with dark brown bars. Dorsum of hands and feet pale brown with dark brown blotches on outer fingers and toes. Flanks pale brown with dark brown markings. Belly, axial region and groin pale orange lacking spots or blotches, pale brown throat finely mottled. The concealed surfaces of limbs are brown finely mottled with dark brown. Ventral surface of tarsus dark brown. Palmar surface pale brown densely mottled with dark brown, subarticular, palmar and thenar tubercles dark brown; dark brown bars originating below the eyes and extending towards the lips ( Fig. 3A–D View FIGURE 3 ).
Measurements of holotype (in mm). SVL 24.4; HL 10.3; HW 10.7; ED 4.4; END 2.1; EW 2.6; IOD 3.2; IND 1.9; ETD 1.2; TYD 1.5; TL 13.7; FL 11.2; FoL 10.8; HaL 7.3.
Males Females
n =17 n =16 Variation. Morphometric variation in adult males and females is described in Table 3. Color variation in life is shown in Figure 5 View FIGURE 5 . Broad dark brown middorsal with narrow cream middorsal lines, and dorsolateral stripes extending from lever of eyelids to posterior end of body (DHMECN 11702, FHGO 8884, 9963, QCAZ 65056, 65063, 65112, 65147). A cream vertebral stripe is present in the holotype and several additional specimens (DHMECN 11702, FHGO 8346, QCAZ 65063, 65098, 64143, 65147). Brown markings on dorsum reduced to the scapular region in a specimen (QCAZ 65096). W-shaped scapular mark on dorsum barely differentiated in the holotype and in some additional specimens (DHMECN 11702, 11693, FHGO 9962, 9965) and strongly defined in a specimen (FHGO 9964). Sacral chevron marks in four specimens (DHMECN 11693, 11697, FHGO 8346, QCAZ 65056, 65063, 65147). Supratympanic stripes barely differentiated in a specimen (QCAZ 65096). Interocular bar is strongly differentiated in a specimen (QCAZ 65110, 65096). Bar lips barely differentiated in two specimens (QCAZ 65096, 65099). Canthal stripe absent in a specimen (QCAZ 65096). A specimen (FHGO 9965) with minute pale brown spots on belly and throat. The row of tarsal tubercles is undistinguished in the preserved specimen FHGO 8264.
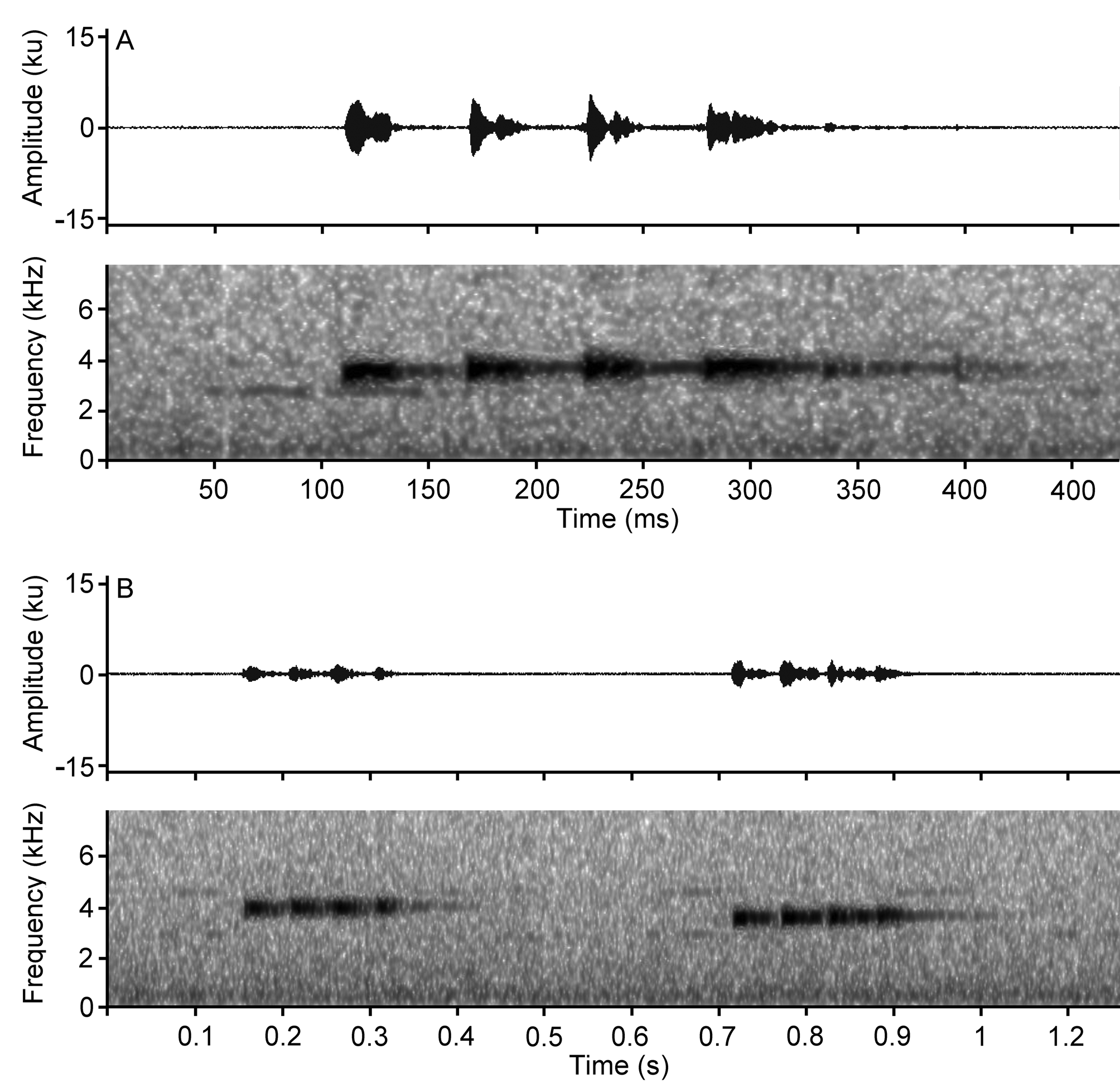
Advertisement call. The advertisement call (mean ± SD is provided for all acoustic parameters analyzed) is characterized by a modulated frequency. The calls are a cadential series of metallic sounds, which could be confused with cricket sounds. Dominant frequency = 3.62 ± 0.10 kHz. Call duration = 222.50 ± 35.64 ms, with intervals = 5393.30 ± 1828.60 ms, Call per minute = 12.27 ± 4.48 calls/minute. Calls are multipulsed by mean = 4.58 ± 0.67 pulses with duration = 17.23 ± 3.24 ms. Calls intervals = 38.80 ± 7.51 ms, producing a mean = 17.93 ± 2.57 pulses/second ( Table 4). Pulses have a slight increase in frequency of 0.02–0.08 kHz between pulses, having 0.12–0.21 kHz between the first and last pulses. Choruses are not orderly structured, males close to each other call antiphonally ( Fig. 6 View FIGURE6 ).
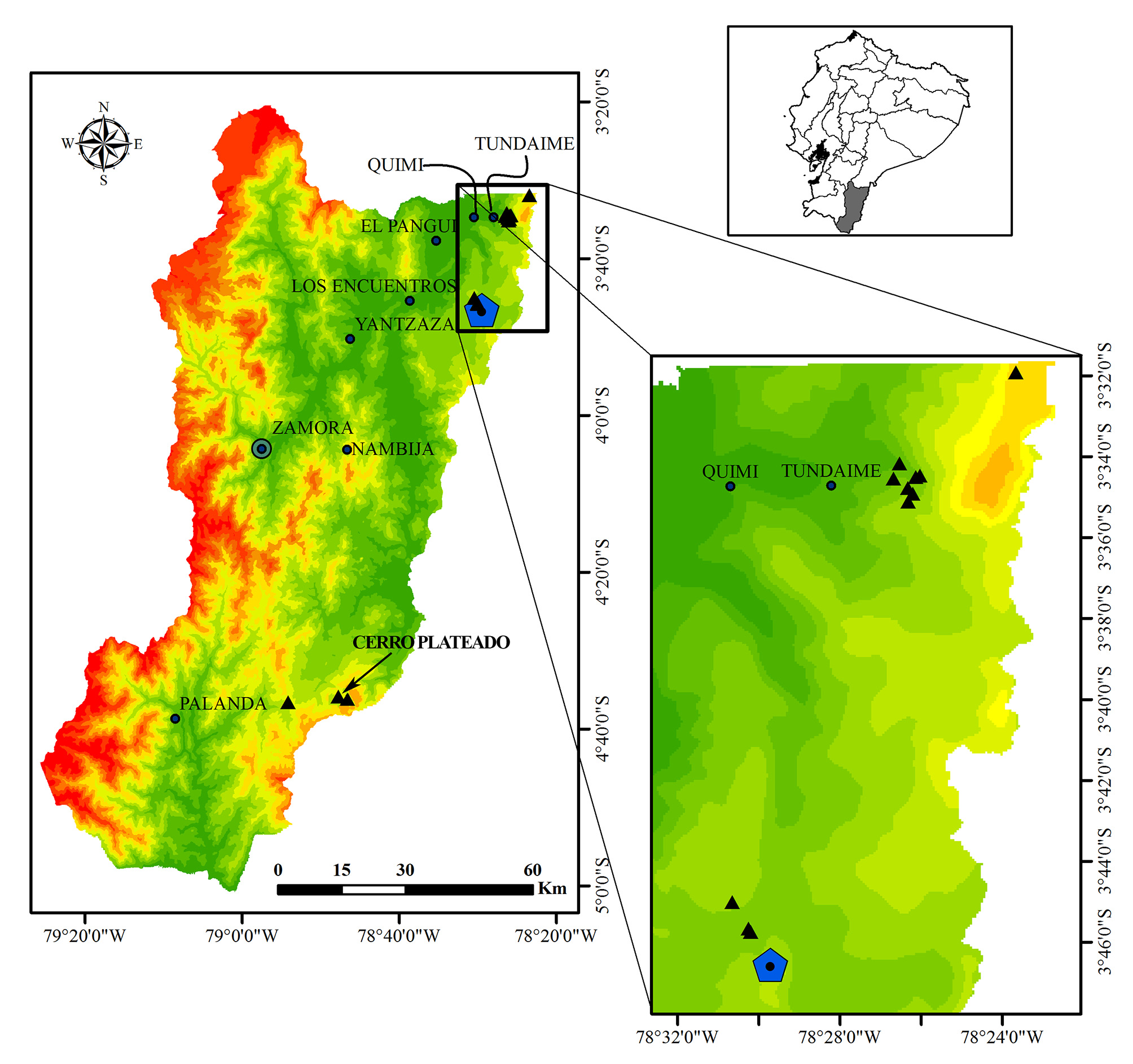
Distribution and natural history. Known only from five localities in the province of Zamora Chinchipe, all in the Cordillera del Condor (La Zarza, Colibrí, Mirador, and Las Peñas), (Cerro Plateado) ( Fig. 7 View FIGURE 7 ). The type locality is in evergreen montane forest between 1200–1800 m elevation (based on Ministerio del Ambiente de Ecuador 2012, ecosystem classification system; Fig. 8A–B View FIGURE 8 ). The following ecological information was obtained at the type locality. Pristimantis yantzaza is nocturnal, males were observed calling partially concealed among leaves. A few males were heard calling very sporadically from other small vegetation islands by day or night, but could not be located. Juveniles were observed on low vegetation or moss, adults on upper leaves of bushes 0.30 to 5.00 m above ground ( Fig. 8C–D View FIGURE 8 ). Two amplectant pairs were collected on a bromeliad ( Guzmania sp.). Sympatric congeners include Pristimantis incomptus , P. prolatus , P. proserpens , and P. serendipitus .
Conservation status. We consider this species to be Endangered following A4B1a,b(iii) IUCN criteria because: (i) its extent of occurrence is estimated to be less than 22 km 2 (i.e., total area of continuous montane forest in the Cordillera del Condor and Andes); (ii) it has been detected in five localities in the province of Zamora Chinchipe, one of which was severely affected by a large scale open-pit mining; and (iii) its habitat is severely fragmented, isolated from other such habitats and declining in extent and quality due to deforestation.




| QCAZ |
Museo de Zoologia, Pontificia Universidad Catolica del Ecuador |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |