
Trilobatus trilobus ( Reuss, 1850 )
|
publication ID |
https://doi.org/10.1080/14772019.2019.1578831 |
|
DOI |
https://doi.org/10.5281/zenodo.10932447 |
|
persistent identifier |
https://treatment.plazi.org/id/072AAD72-2332-AE7F-3BDC-FACCFE57F2AA |
|
treatment provided by |
Felipe |
|
scientific name |
Trilobatus trilobus ( Reuss, 1850 ) |
| status |
|
Trilobatus trilobus ( Reuss, 1850) View in CoL
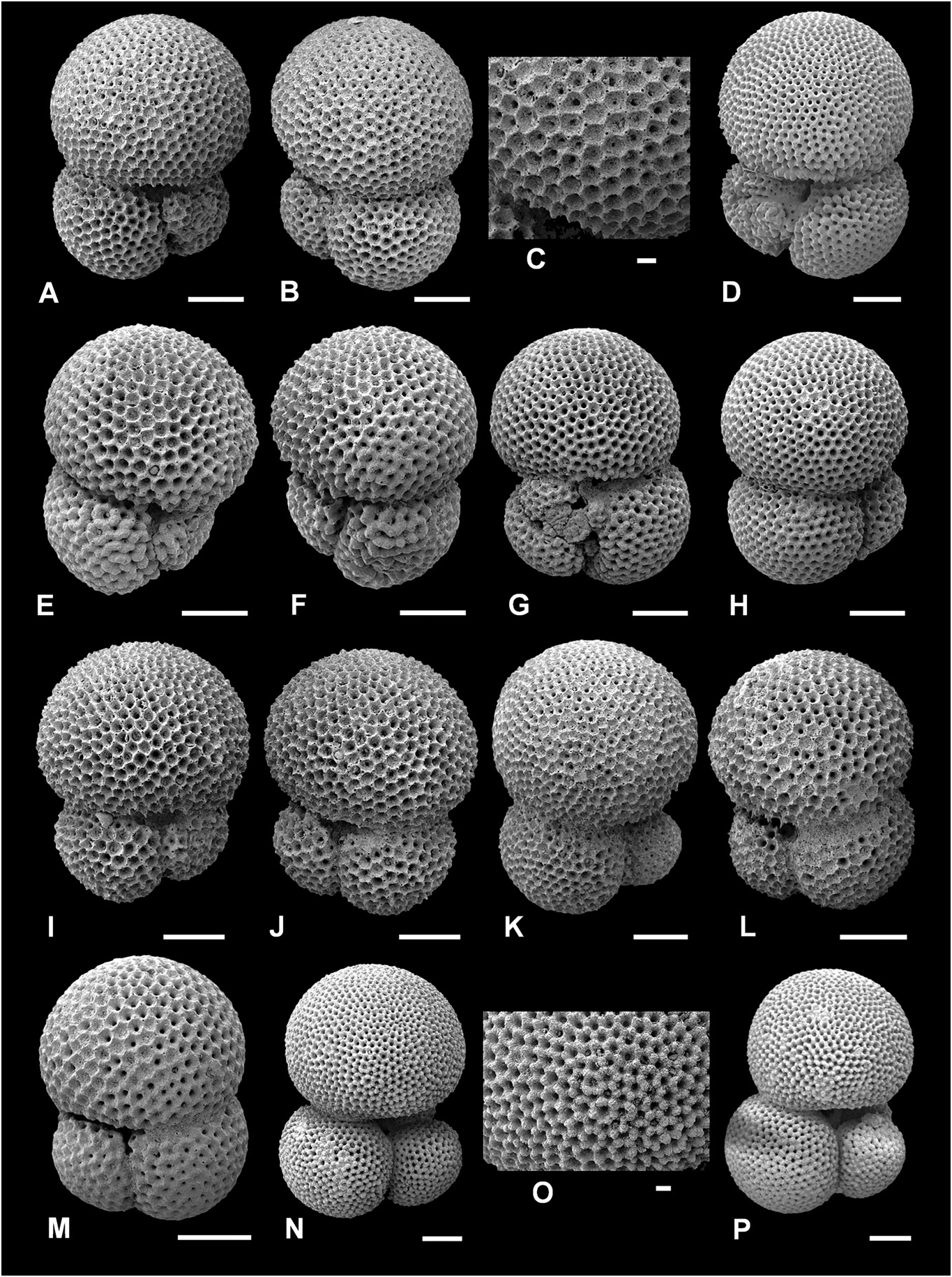
( Figs 6A–O View Figure 6 , 16E, 17A, E View Figure 17 )
1850 Globigerina triloba Reuss : 374, pl. 47, fig. 11a–e.
1957 Globigerinoides triloba triloba (Reuss) ; Bolli: 112, pl. 25, fig. 2a–c.
1960 Globigerinoides triloba triloba (Reuss) ; Jenkins: 353, pl. 2, fig. 5a–c.
1966 Globigerinoides trilobus trilobus (Reuss) ; Jenkins: 9, pl. 2, fig. 8a–c.
1967 Globigerinoides quadrilobatus trilobus (Reuss) Closs : 340, pl. 1, fig. 22.
1975 Globigerinoides quadrilobatus trilobus (Reuss) Srinivasan : 139, pl. 2, fig. 7.
1983 Globigerinoides triloba (Reuss) Kennett & Srinivasan : 62, pl. 13, figs 1–3.
1994 Globigerinoides trilobus (Reuss) Loeblich & Tappan : 107, pl. 206, figs 1–6.
2012 Globigerinoides trilobus (Reuss) Rögl : 181, pl. 1, figs 1–7.
2018 Trilobatus trilobus (Reuss) Spezzaferri, Olsson, & Hemleben : 300–302, pl. 9.14, figs 1–21.
Description. Type of wall: spinose, coarsely cancellate (often termed polygonal or honeycomb) ‘ sacculifer - type’ wall texture. Test morphology: low trochospire, initially involute, later more evolute coiling, coiling direction random, three globose, near-spherical chambers in the final whorl, increasing rapidly in size as added, final chamber larger than all other chambers combined; sutures distinct, depressed, straight to slightly curved on both sides; umbilicus narrow; primary aperture interiomarginal-umbilical, a low arch, slit-like, no bordering rim; supplementary apertures small, placed at the sutures of the preceding chamber and third-previous chamber, often only one visible due to infilling or secondary calcification.
Note: description is based on the original description and species concept of Reuss (1850, p. 374), and also Kennett & Srinivasan (1983, p. 62) and Rogl (2012, p. 182), but is here emended and extended.
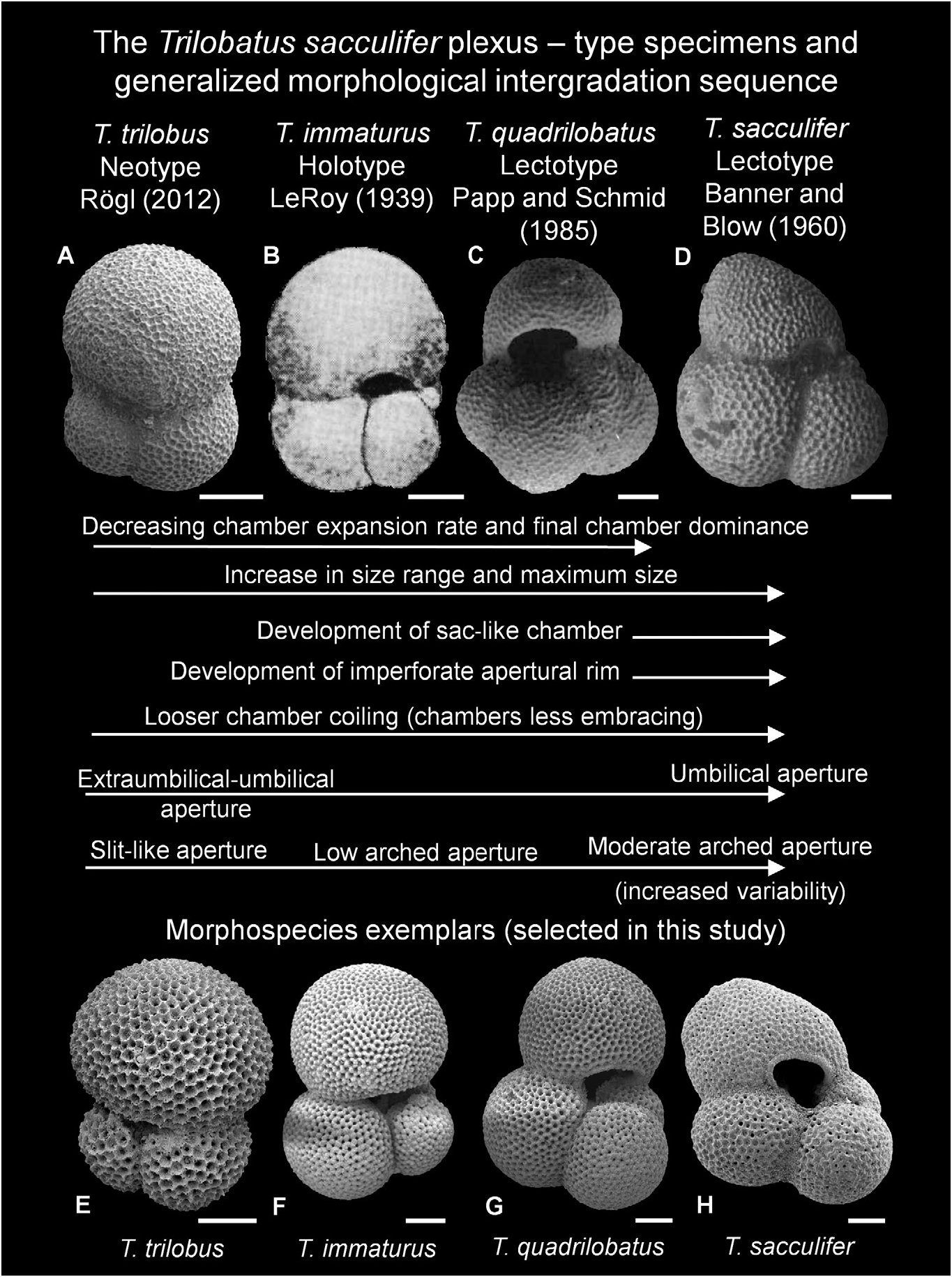
Remarks. Trilobatus trilobus is distinguished from T. immaturus and T. quadrilobatus by having more rapidly enlarging chambers in the final whorl, resulting in a final chamber that is larger than all other chambers combined. The last chamber is more embracing of the earlier chambers owing to tighter coiling, whereas T. immaturus and T. quadrilobatus are more loosely coiled with generally more than three chambers in the final whorl (3.5 to 4). Trilobatus trilobus differs from T. sacculifer which possesses a sac-like final chamber and from G. fistulosa which has protuberances. Globigerinoides altiaperturus Bolli, 1957 differs primarily in possessing a high-arched, semi-circular primary aperture, whereas in T. trilobus it is low and slit-like.
Type locality. Reuss (1850, p. 374) first described Globigerina triloba , documenting its occurrence in five separate localities from four countries ( Romania, Poland, Austria [two localities] and Italy; see also Rogl 2012, p. 181 for details). Rogl (2012) illustrated numerous specimens from these localities and designated a neotype (here reproduced in Fig. 17 View Figure 17 ) from the Polish locality material of Reuss (1867). Therefore, the type locality is the salt mine Wieliczka, near Krakow, Poland.
Taxonomic history. Reuss (1850, p. 374) described the new species Globigerina triloba and illustrated a specimen that clearly exhibits multiple apertures (pl. 47, fig. 11a–e). Reuss (1850) also highlighted how the last chamber is larger than the preceding chambers. The sutures are deeply incised, giving this morphospecies a distinct ‘three-lobed’ appearance, which explains the derivation of the name trilobus . After the introduction of Globigerinoides Cushman, 1927 , T. trilobus was accordingly assigned to this genus by later workers (see synonymy list).
Bolli (1957, p. 113) named Globigerinoides triloba altiapertura (= Globigerinoides altiaperturus ) as a subspecies for forms similar to T. trilobus and T. immaturus morphotypes, but possessing a higher arched, semi-circular primary aperture (see also Bolli & Saunders 1985, p. 192). However, G. altiaperturus is probably not closely related to T. trilobus (see Kennett & Srinivasan 1983, text-fig. 9; and Spezzaferri et al. 2015, fig. 5). Whilst G. altiaperturus is included in the above distinguishing features section, these morphospecies should not be confused morphologically due to their contrasting primary aperture morphology and wall texture.
Trilobatus trilobus has the most conservative morphology of the morphospecies in this study, as exemplified in Figure 6 View Figure 6 . This is mostly attributable to our strict concept of T. trilobus regarding the dominance of the final chamber; it must be larger than all of the preceding chambers combined ( sensu Bolli 1957) (see Morphometrics and biometrics). Trilobatus trilobus intergrades with T. immaturus , so using the relative size of the last chamber as the primary delimiting character is arbitrary. However, it can be estimated without using scanning electron microscopy and is easily quantified using morphometrics, thus making it an excellent discerning feature. In the morphometric results, a T. trilobus final chamber dominance ratio ( FCDR) must be greater than or equal to 1, whereas in T. immaturus the FCDR value must be less than 1 ( Fig. 19 View Figure 17 ). The ratio compares chamber area (Lm 2) from a two-dimensional image (see Methods). In T. trilobus , the final chamber volume should also be larger than the rest of the test volume. As the final chambers always have a regular, globose morphology (i.e. no flattening or extension as in T. sacculifer ), specimens with ratios of more than 1 can also be used to equate to a larger chamber volume (Lm 3) ratio. Typically, the maximum diameter of the final chamber is also greater than the maximum diameter of the rest of the test. However, occasionally the first two chambers of the final whorl appear marginally wider than the last chamber (e.g. Fig. 6D View Figure 6 ), yet this is still considered a T. trilobus because of the dominant last chamber (i.e. an FCDR value of> 1).
Conversely, if the final chamber is not larger than all preceding chambers, then the specimen cannot be T. trilobus . Essentially, this means that T. trilobus cannot possess a kummerform final chamber ( sensu Berger 1969; see also Olsson 1973). This consequently results in many specimens with kummerform final chambers being assigned to T. immaturus (e.g. Fig. 7A–D View Figure 7 ; see T. immaturus , Remarks).
Trilobatus trilobus has a sacculifer - type wall texture ( Fig. 6C View Figure 6 ), although this is commonly obscured by secondary, ‘gametogenic’ calcite. In particular, the initial chambers of the final whorl are more heavily calcified and the sacculifer - type wall texture is less evident (e.g. Fig. 6C View Figure 6 ). Secondary calcification also affects the preceding whorl on the spiral side, and the small chambers are often indistinct. If the preceding whorl cannot be clearly observed to identify the spiral side, it is distinguished by the position of the largest aperture. The umbilical side possesses an extraumbilical-umbilical aperture, whereas the spiral side has a central supplementary aperture placed at the base of the final chamber at the sutures of the preceding and third-preceding chambers.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Globigerinacea |
|
Family |
|
|
SubFamily |
Globigerininae |
|
Genus |