
Trilobatus immaturus ( LeRoy, 1939 )
|
publication ID |
https://doi.org/10.1080/14772019.2019.1578831 |
|
DOI |
https://doi.org/10.5281/zenodo.10932455 |
|
persistent identifier |
https://treatment.plazi.org/id/072AAD72-2336-AE63-3918-FDEDFD90F5AE |
|
treatment provided by |
Felipe |
|
scientific name |
Trilobatus immaturus ( LeRoy, 1939 ) |
| status |
|
Trilobatus immaturus ( LeRoy, 1939) View in CoL
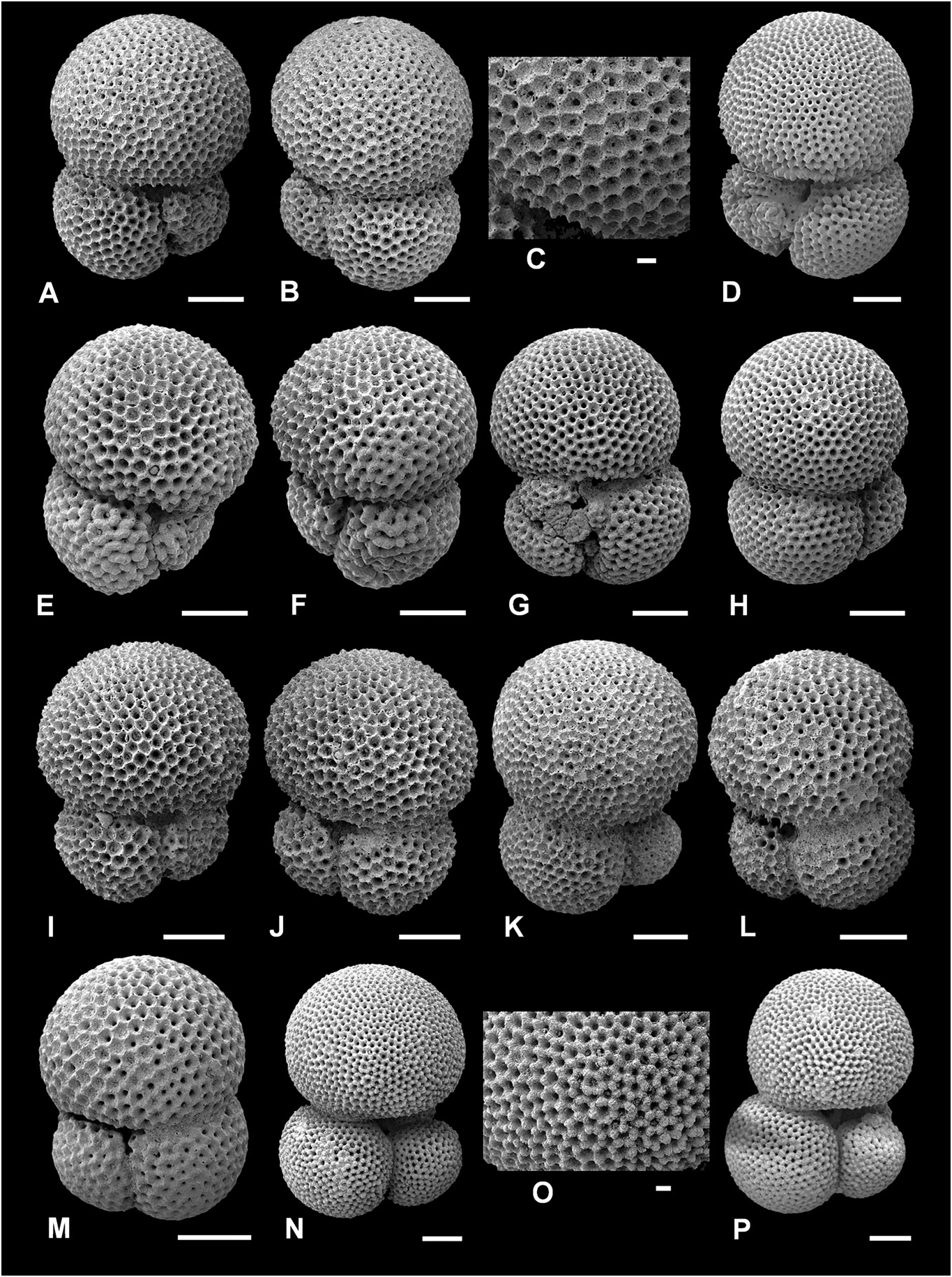
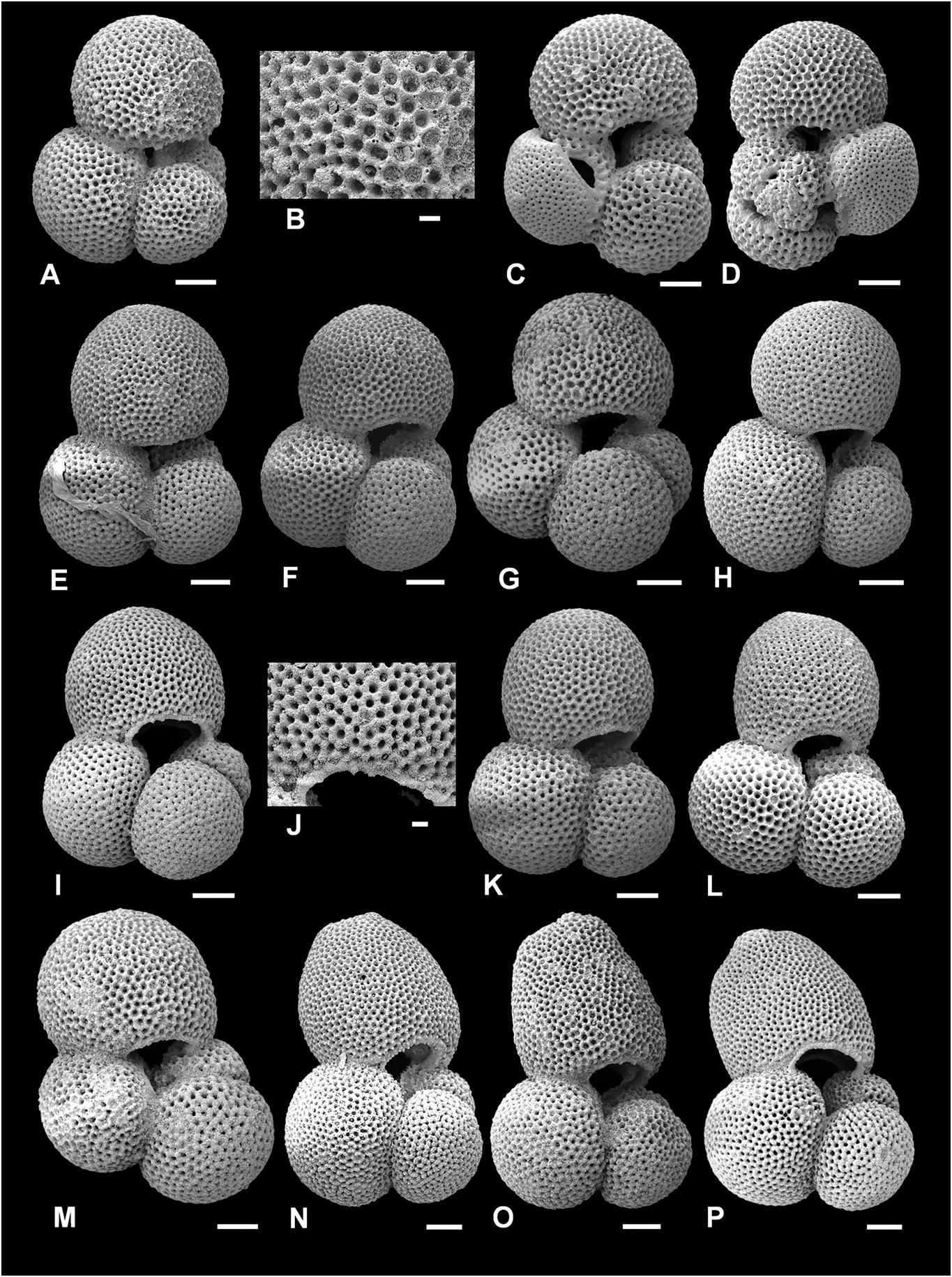
( Figs 6P View Figure 6 , 7A–K View Figure 7 , 17B, F View Figure 17 )
1939 Globigerinoides sacculifera var. immatura LeRoy : 263, pl. 3, figs 19–21.
1957 Globigerinoides triloba immatura LeRoy ; Bolli: 113, pl. 25, figs 3a–4c.
1964 Globigerinoides triloba immatura LeRoy ; LeRoy: F42, pl. 14, fig. 16.
1967 Globigerinoides quadrilobatus immaturus LeRoy ; Closs: 340, pl. 1, fig. 15.
1969 Globigerinoides triloba immatura LeRoy ; Mohan: 36, pl. 4, figs 3, 4.
1975 Globigerinoides quadrilobatus immaturus LeRoy ; Srinivasan: 138, pl. 2, fig. 4.
1981 Globigerinoides quadrilobatus immaturus LeRoy ; Chaproniere: 112, fig. 5D, a–d.
1983 Globigerinoides immaturus LeRoy ; Kennett & Srinivasan: 64, pl. 13, figs 7–9.
2018 Trilobatus immaturus LeRoy ; Spezzaferri, Olsson, & Hemleben: 289–292, pl. 9.9, figs 1–21.
Description. Type of wall: spinose, coarsely cancellate (often termed polygonal or honeycomb) ‘ sacculifer - type’ wall texture. Test morphology: low trochospire, 3 to 3.5 globose, spherical chambers in the final whorl increasing moderately in size as added; sutures distinct, depressed, straight to slightly curved on both sides; umbilicus narrow; primary aperture extraumbilical-umbilical, generally a low arch, typically no bordering rim; small supplementary apertures on spiral side, one per chamber, placed at the sutures of the preceding chamber and third-previous chamber, sometimes the only one visible due to infilling or secondary calcification.
Note: description is based on the original species concept of LeRoy (1939, p. 263) and also Bolli (1957, pp. 112–113) and Kennett & Srinivasan (1983, p. 64), but is here emended and extended.
Remarks. Trilobatus immaturus is distinguished from T. trilobus by having looser coiling and a lower rate of chamber expansion in the final whorl, resulting in a final chamber that is not larger than the other chambers combined. Trilobatus quadrilobatus differs in typically possessing four chambers in the final whorl, and having a lower rate of chamber expansion, resulting in almost equal-sized chambers. The last chamber is more embracing of the earlier chambers owing to tighter coiling, whereas T. quadrilobatus is more loosely coiled. Thus, T. immaturus can be seen as intermediate in morphology between T. trilobus and T. quadrilobatus . Trilobatus immaturus differs from T. sacculifer which possesses a sac-like final chamber and G. fistulosa which has protuberances.
Type locality. Telisa Shales , Tapoeng Kiri area, Rokan-Tapanoeli, central Sumatra, Indonesia .
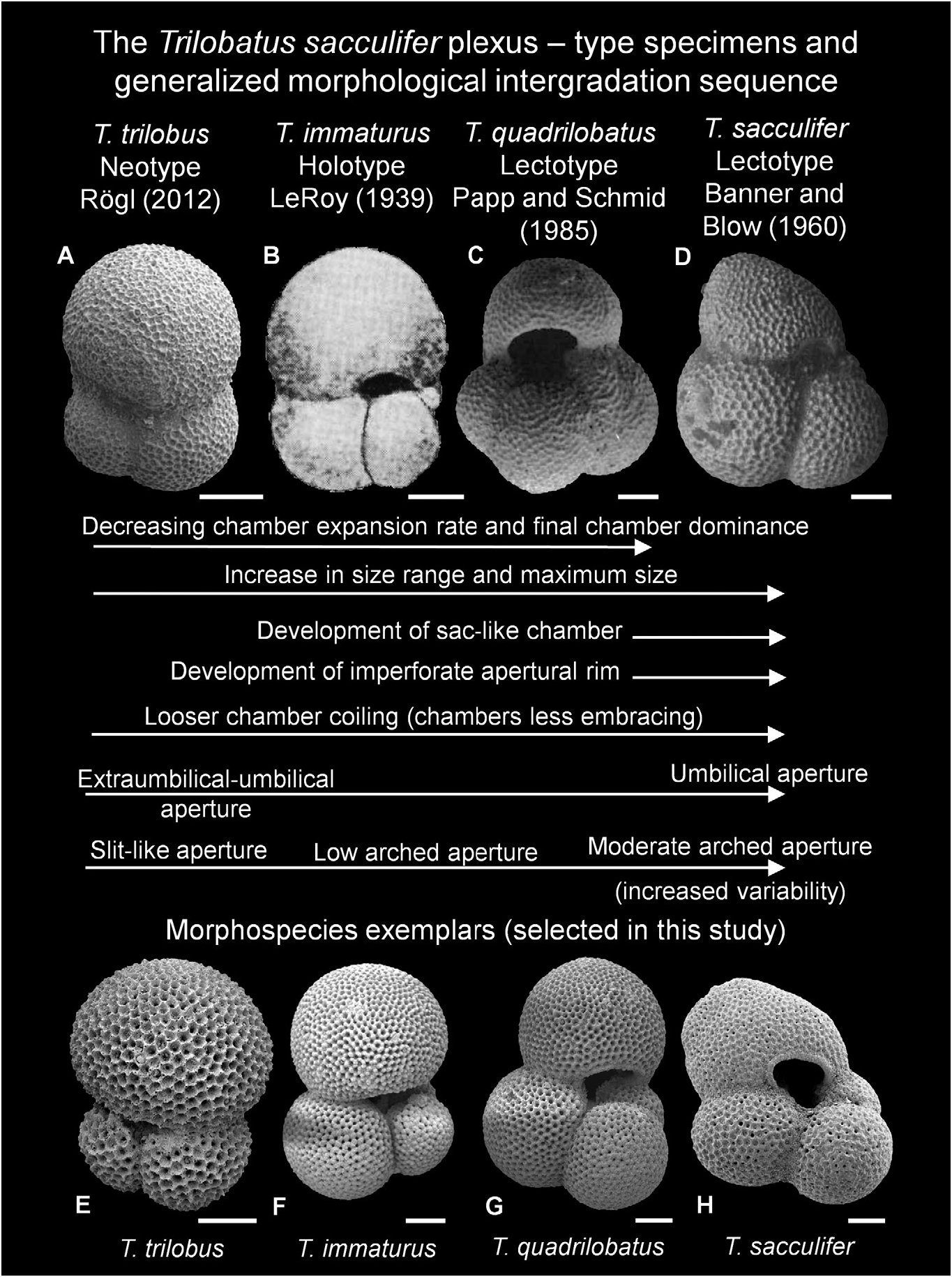
Taxonomic history. Trilobatus immaturus was named by LeRoy (1939, pp. 263–264) as Globigerinoides sacculiferus var. immatura . LeRoy (1939) originally considered Globigerinoides sacculiferus var. immatura a varietal form of T. sacculifer and thus did not provide a formal description. He noted the close morphological similarity with T. sacculifer but considered it “either an early or immature form” ( LeRoy 1939, p. 263) because of the absence of the characteristic sac-like final chamber, and this explicates the derivation of the name. The immaturus variety was assigned to Globigerinoides Cushman, 1927 , based on the presence of multiple apertures which LeRoy (1939) clearly illustrated in the holotype images. These illustrations depict a morphotype broadly similar to that of the original images for T. trilobus by Reuss (1850), but it has slightly more than three chambers (approximately 3.5) and a slightly lower chamber expansion rate in the final whorl (see Fig. 17 View Figure 17 ). Bolli & Saunders (1985) suggested that the differences were minimal; however, Bolli’ s (1957) concept is well suited to delimit the morphospecies.
Bolli (1957, p. 112) clarified the differences between T. trilobus and T. immaturus , stating that “ G. triloba triloba differs from G. triloba immatura in having a final chamber that is larger than all the earlier chambers combined” (see also Jenkins [1960, p. 354] and Kennett & Srinivasan [1983, p. 64] for their comparable distinctions). These differences were also well illustrated by Bolli (1957, i.e. compare text-fig. 21.1 and pl. 25, fig. 2a–c [ T. trilobus ] with text-fig. 21.2 and pl. 25, figs 3a–4c [ T. immaturus ]). Although originally naming immaturus as a varietal form of sacculifer, LeRoy (1964) subsequently followed Bolli’ s (1957) designation of immaturus as a subspecies of trilobus . In contrast, other workers (e.g. Closs 1967; Srinivasan 1975; Chaproniere 1981) considered T. immaturus to be a subspecies of quadrilobatus (see synonymy list), following Blow & Banner’ s (1960, 1965) assertion regarding the aforementioned senior priority of quadrilobatus and it being a ‘central form’. Trilobatus immaturus also has an intermediate morphology between T. trilobus and T. quadrilobatus , and intergrades with both ( Figs 6–8 View Figure 6 View Figure 7 View Figure 8 ), and thus is often reported as a subspecies of T. trilobus and T. quadrilobatus .
Kennett & Srinivasan (1983) avoided a trinomial nomenclature for this group, giving T. immaturus species status (as Globigerinoides immaturus ), as binomial nomenclature is now standard taxonomic practice for planktonic foraminifera (e.g. Olsson et al. 1999; Pearson et al. 2006; Pearson & Wade 2015; Wade et al. 2018a). Since the taxonomic revision of Spezzaferri et al. (2015), immaturus has been assigned to the newly erected genus Trilobatus , and we adhere to this designation.
In some respects, T. immaturus can be considered a ‘wastebasket’ taxon (e.g. Longoria & Gamper 1995, p. 332; Aze et al. 2011, p. 921; Pearson & Wade 2015, p. 12), although note the slightly different meaning envisaged herein. Firstly, owing to the strict morphological concept of T. trilobus , that morphospecies cannot possess a kummerform final chamber (see T. trilobus Taxonomic history, above). This means that T. immaturus has a high proportion of forms with a kummerform final chamber (see Fig. 7A, B View Figure 7 ), because it is augmented by the inclusion of forms which cannot be referred to as T. trilobus . This may also explain why the maximum size of T. immaturus is often similar to that of T. trilobus ( Fig. 18 View Figure 18 ), rather than being more of an intermediate size between T. quadrilobatus and T. trilobus . Secondly, whilst ‘typical’ T. trilobus and T. quadrilobatus have distinctive morphology, the name T. immaturus has come to represent any intermediate morphology which does not fit with the T. trilobus and T. quadrilobatus morphospecies concepts. Nevertheless, Andŕe et al. (2013, fig. 8) highlighted that the morphospecies name immaturus has seen the rarest usage of the T. sacculifer plexus names in the fossil record to Recent, in comparison with the three-chambered T. trilobus and the three and a half- to four-chambered T. quadrilobatus .
The wall texture is sacculifer - type (exemplified in Fig. 7C, D View Figure 7 ), although as with all T. sacculifer plexus members, secondary calcification commonly obscures the original wall texture and especially the spine holes ( Fig. 7J, K View Figure 7 ). The secondary calcification does not affect all chambers equally; the close-up images in Figure 7J and K View Figure 7 are from the final and penultimate chambers, respectively. The penultimate chamber has probably been affected more severely by secondary calcification, and also previous chambers often add more calcite during precipitation of succeeding chambers.
Trilobatus sacculifer is generally considered to have arisen from T. quadrilobatus (e.g. Kennett & Srinivasan 1983). However, we also observe intergradation between T. immaturus and T. sacculifer ( Fig. 7B–H View Figure 7 ), despite T. immaturus usually being intermediate in morphology between T. trilobus and T. quadrilobatus . These specimens show clear evidence of imperforate lips on the primary and first supplementary apertures, and the final chambers are slightly more flattened than ‘typical’ T. immaturus specimens. Though the T. immaturus - T. sacculifer intermediate morphotypes ( Fig. 7B–H View Figure 7 ) occur less frequently than T. quadrilobatus - T. sacculifer intermediate morphotypes ( Fig. 8H–N View Figure 8 ), a sac-like chamber may also develop on forms which would otherwise be referred to T. trilobus , highlighting how the morphospecies are closely related. The fossil evidence presented here corresponds with the evidence from Recent forms which suggests that the morphospecies of the T. sacculifer plexus represent one (biological) species, as the sac-like chamber may develop from T. trilobus , T. immaturus or T. quadrilobatus .


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Globigerinacea |
|
Family |
|
|
SubFamily |
Globigerininae |
|
Genus |