
Liolaemus caparensis, Breitman, Maria Florencia, Pérez, Cristian Hernán Fulvio, Parra, Micaela, Morando, Mariana, Sites, Jack Walter & Avila, Luciano Javier, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.204780 |
|
DOI |
https://doi.org/10.5281/zenodo.6191226 |
|
persistent identifier |
https://treatment.plazi.org/id/073487A0-FFC1-FE34-FF24-C974FCA5FF06 |
|
treatment provided by |
Plazi |
|
scientific name |
Liolaemus caparensis |
| status |
sp. nov. |
Liolaemus caparensis sp. nov.
( Figure 2 View FIGURE 2 )
Holotype. MLP.S 2628 ( Figure 2 View FIGURE 2 ), an adult male from Campo Las Piedras plateau, 7 km N Punta del Lago ranch, Lago Argentino department, Santa Cruz province, Argentina (49º 34’ 11.0” S, 72º 02’ 51.9” W, 868 m; Figures 3 View FIGURE 3 , 4 View FIGURE 4 ), L.J. Avila, C.H.F. Pérez, M.F. Breitman and N. Feltrin collectors, 16th January 2008.
Paratypes. LJAMM –CNP 9379–9380 adult males, LJAMM –CNP 9383–9387 females and LJAMM –CNP 9382, 9388, 9389 juveniles; from the same locality of holotype, L.J. Avila, C.H.F. Pérez, M.F. Breitman and N. Feltrin collectors, 16 th January 2008.
Diagnosis. Liolaemus caparensis sp. nov. is a member of the magellanicus group, in the L. lineomaculatus section, based on molecular evidence ( Figure 1 View FIGURE 1 ). Liolaemus caparensis sp. nov. differs from L. magellanicus in the following traits: shorter tibia-fibula length (6.19-8.62, X = 7.24 vs. 6.91-10.76, X = 9.09; p <0.0001), shorter radius-ulna length (5.24-5.66, X = 5.24 vs. 5.22-6.88, X = 5.98; p = 0.0017), larger number of midbody scales (39- 55, X = 43.82 vs. 34-48, X = 40.44; p = 0.0282), larger number of dorsal scales (39-47, X = 41.45 vs. 34-45, X = 37.5; p = 0.0003), more ventral scales (64-73, X = 68.36 vs. 49-77, X = 59.17; p <0.0001), more infradigital lamellae of the third finger (14-18, X = 15 vs. 11-15, X = 13.33; p = 0.0006), and more infradigital lamellae of the fourth toe (18-23, X = 20.3 vs. 15-20, X = 18.24; p = 0.0011) (for details see Tables 1 View TABLE 1 to 4). Liolaemus caparensis sp. nov. has smaller dorsal blotches on the hindlimbs and a more defined vertebral line in comparison with L. magellanicus ( Figure 5 View FIGURE 5 ), as well as fewer precloacal pores than L. magellanicus (3-4 vs. 3-5). Dorsal scales of Liolaemus caparensis sp. nov. have a shorter mucron than those of L. magellanicus . The dorsal surface of limb scales is more carinated and less mucronated in L. caparensis sp. nov. than in L. magellanicus ( Figure 5 View FIGURE 5 ).
Description of holotype. Adult male. Snout-vent length 59.0 mm. Tail length (complete, not regenerated) 84.0 mm. Radius-ulna length 25.9 mm. Head length 13.4 mm (from anterior border of tympanum to tip of snout), 11.0 mm wide (at anterior border of tympanum), 7.7 mm long (at anterior border of tympanum). Snout length 4.9 mm (orbit-tip of snout distance). Auditory meatus-eye distance 4.7 mm. Interorbital distance 4.2 mm. Eye-nostril distance 3.1 mm. Forelimb length 17.5 mm. Tibial length 8.6 mm. Foot length 14.6 mm (ankle to tip of claw on fourth toe).
Dorsal head scales slightly bulged, smooth, 14 between occiput, at level of anterior border of tympanum, to rostral, pitted with numerous scale organs in the anterior region, and reducing to no organ in the posterior half of the head. Rostral scale wider (2.8 mm) than long (0.9 mm). Two postrostrals that together with anterior lorilabial separate nasal scales from rostral, surrounded by six scales. Nasal scales longer than wide, irregularly hexagonal; nostril one-half length of nasal, posterior in position. Scales surrounding nasals 7 on each side. Four internasals. Frontonasals and prefrontals 14, irregular in shape, size and position. Two frontal scales. Frontoparietals in two rows, two anterior and one posterior scale. Interparietal pentagonal (1.3 mm), surrounded by five scales; three smaller and irregular in front and sides, two larger in back. Parietal eye evident. Parietals slightly bulged, irregularly shaped, equal in size to interparietal. Circumorbitals: 9 on the left side, incomplete; 10 on the right side, incomplete. Transversally expanded supraoculars 4–3. Smaller lateral supraoculars: 16–19. Three canthals on the left side, one posterior large, higher than wide, in contact with postnasal, two inferior, small; four canthal on the right side, one posterior with similar characteristics that the left side, and three inferior, small. Loreal scales slightly bulged, 4–5 on each side. Lorilabials longer than wide (7–8), slightly smaller than labials. Superciliaries 7 on each side, flattened and elongated, anterior four broadly overlapping dorsally. Orbit with 13–12 upper and 11–10 lower ciliaries on each side. Orbit diameter 4.7 x 2.3 mm. Preocular small, unfragmented, longer than wide. Subocular scale elongated, approximately nine times longer than wide (4.0 x 0.5 mm). A well marked longitudinal ridge along upper margin of preocular and subocular scales. Postocular small, slightly bulged, quarter superimposed to subocular, with a marked longitudinal ridge. Palpebral scales, small granular and bulged. Supralabials 6 on each side, convex. Temporals smooth, convex, juxtaposed with one scale organ in the tip. Anterior auriculars smaller than adjacent posterior temporals, slightly projecting outward (1 on each side). Posterior auriculars small and granular. External auditory meatus conspicuous, rounded (1.8 x 1.5 mm). Lateral scales of neck granular with inflated skin. Mental scale wider (3.0 mm) than high (1.3 mm), in contact with four scales. Mental followed posteriorly by two postmentals, and two rows of 2–1 chinshields. Five infralabials on each side, first on each side quadrangular two times wider than supralabials, all others elongated, slightly smaller than supralabials. Gular scales smooth, flat, imbricate, with rounded posterior margins, with melanophores. Scales of throat between chinshields slightly imbricate. Twenty-six gular scales between tympanum openings. Infralabials separated from chinshields by one to two rows of scales. Antehumeral and longitudinal neck folds well developed; gular fold incomplete; postauricular, rictal, dorsolateral and oblique folds unconspicuous.
Scales of dorsal neck region rhomboidal, imbricate, strongly keeled, mucronate. Thirty-nine scales between occiput and anterior surface of thighs. Dorsal body scales rhomboidal, imbricate, strongly keeled and mucronate. Dorsal scales grade laterally into slightly smaller, slightly keeled scales at midbody. Scales immediately anterior and posterior to forelimb and hindlimb insertion small, smooth, granular, and non-overlapping. Body lateral scales grading smaller to larger at midbody. Ventral body scales rhomboidal, smooth, flat, imbricate, and bigger than dorsal scales. Forty-four midbody scales; scales between mental and precloacal pores 71. Scales of cloacal region about equal in size to ventral body scales. Four precloacal pores.
Anterior suprabrachials rhomboidal, imbricate, smooth, slightly larger in size to dorsal body scales. Postabrachials smaller, smooth, becoming granular near axilla. Supra-antebrachials rhomboidal, imbricate, smooth, some of the external lateral slightly keeled. Infra-antebrachials rhomboidals, imbricate, smooth, toward the hand slightly keeled. Supracarpals imbricated, rhomboidal, smooth. Infracarpals strongly imbricate, rhomboidal, slightly keeled and mucronate. Infradigital lamellae with 3-keels, each terminating in a short mucron, 3-mucronate, numbering: I: 8, II: 12, III: 16, IV: 19, V: 11. Claws robust, curved and sharp, opaque brown.
Suprafemorals as large as dorsal body scales, rhomboidal, imbricated, strongly keeled and mucronate. Postfemorals small, granular shape. Supratibial rhomboidal, imbricated, strongly keeled and mucronate. Infrafemoral scales small, granular and smooth. Supratarsals rhomboidals, imbricated and smooth, some lateral scales slightly keeled. Infratarsals small, rhomboidal, imbricate, smooth, mucronate, some slightly keeled and mucronate near the digit. Subdigital scales 3-keeled, 3-mucronate, numbering: I: 9, II: 13, III: 19, IV: 21, V: 14. Claws robust, curved and sharp, opaque brown. Tail complete, non-regenerated. Dorsal and lateral caudal scales, rhomboidal in the first half of the tail, becoming quadrangular toward the tip, strongly keeled and mucronate. Ventrals subtriangular and smooth, toward posterior half moderately keeled.
Color of holotype in life. Grey dorsal background ( Figure 2 View FIGURE 2 ). The posterior borders of the dorsal scales, between neck and pelvic region, present one or two contrasting light colors (white or light blue). Dorsal pattern with eight series of paravertebral and quadrangular black (with a white end) blotches, appearing from the nuchal region to about the first autotomy line, those series fuse into a dark line that is present to the tip of the tail. Dorsolateral region, between the occipital and the pelvic regions, has eight irregular black and white-bordered blotches. Between the series of blotches there are three longitudinal lines, a white vertebral and two light brownish paravertebral lines, the last ones are wider than the vertebral (one and half scale vs. a quarter of scale). The coloration in the vertebral and lateral area is light brown.
Anterior region of the head shows a dark gray background with a central black blotch from the internasals to the frontal scales. Orbitals, postfrontals and parietals are light brown, the black circumorbital scales continue in a line of the same color until the nuchal region. Gray temporal and lateral head regions. Limbs dorsal region are gray with a scarce black reticulation with some lighter color scales.
Ventral scales with melanophores. Black color is present throughout all the ventral body until the postcloacal region. Dark reticulate area is present in the throat and extends to the adjacent malar and maxilla region. Cloacal and postcloacal regions are black. Ventral area of lower belly and femoral region present a bright yellow color. Tail is light gray ( Figure 2 View FIGURE 2 ).
Color of holotype in preservative. After three years in preservative, the dorsal coloration of the head, dorsum, body flanks and tail becomes darker while maintaining the contrast, but the two light brownish paravertebral lines turned gray. Ventral scales of throat, neck, chest, belly and forelimbs maintain the same dark coloration as in life, and the distinctive yellow ventral coloration of the femoral and belly regions turns gray ( Figure 5 View FIGURE 5 ).
Variation. Morphological and meristic characters variation, between males and females of Liolaemus caparensis sp. nov. are shown in Table 5 View TABLE 5 . Females in life present basically the same dorsal and lateral pattern as present in males, but the coloration has a light brown background; vertebral and paravertebral lines are wider and much more notorious than the ones in males, the lines are white; although, one of the lizards presents yellowish dorsolateral lines. Only the females present a white lateral line from the axilla to the groin. Ventral coloration shows a dark reticulate pattern in throat, melanic chest and belly, but two specimens present a yellowish coloration in the ventrolateral, femoral, cloacal and postcloacal regions. All distinctive dorsal lines, and ventral coloration, femoral and lower belly areas turns in preservative from yellowish or bright yellow to gray.
Etymology. The specific epithet of this species “ caparensis ” refers to the name “Capar” given to the actual Viedma lake by the Aónikenk aborigines that inhabited this area. The Campo Las Piedras plateau is located on the northwestern edge of Viedma Lake.
Distribution. Liolaemus caparensis sp. nov. is known only from the type locality, from Campo Las Piedras plateau, 7 km N Punta del Lago ranch, Lago Argentino department, Santa Cruz province, Argentina. 868 m (Figures 3 and 4).
Natural history. The new species is viviparous and probably insectivorous (inferred from the biology of its sister species Liolaemus magellanicus ; Ibargüengoytía et al. 2010). Out of five females, only two presented a ventral yellowish coloration and one of those was gravid, thus we inferred that the yellowish coloration in females is not directly related with the reproductive state. The specimens were found in the Subantarctic Phytogeographic province, Estepa de Festuca pallescens district, in an environment mainly characterized as Festuca grasslands ( Roig 1998). Other dominant vegetation included short bushes (Anartrophyllum desideratum, Senecio spp.) and grasses ( Stipa spp.) ( Figure 3 View FIGURE 3 ). This species was found in open substrates between rocks, sharing the habitat with Liolaemus tari .
Concluding remarks. Lizards from the magellanicus group have a general phenotype (general body form, coloration, and color pattern) similar to lizards of the lineomaculatus group, but they have precloacal pores, a synapomorphy shared with all other species of the Liolaemus lineomaculatus section ( kingii + archeforus and somuncurae groups) but not with species from the lineomaculatus group.
Fouquet et al. (2007), proposed for Neotropical frogs a mtDNA approach to species delimitation based on isolation-by-distance population structure. The method tests for correlation of geographic distance with genetic distance, which characterized most samples up to uncorrelated values of 3%. At this value the isolation-by-distance correlation was not significant and Fouquet et al. (2007) interpreted this break as the limit to intra-specific gene flow.
Martinez (personal communication) found that within Liolaemus the mean genetic distance value of cytochrome b between sister species was ~ 4%; moreover, she found that different groups of Liolaemus had values ranging from 1 to 6%. In the L. lineomaculatus section, average genetic distance between described species is 1.6% (based on the sister species L. kolengh vs. L. hatcheri ), and 2.25% between species of the kingii + archeforus group (based on sister species L. archeforus vs. L. chacabucoense and L. escarchadosi vs. L. tari ). Even though the genetic distance values calculated for L. caparensis sp. nov. relative to L. magellanicus are slightly higher than values of other species from the L. lineomaculatus section, we want to highlight that the discovery of the new species started with the genetics analyses presented by Breitman et al. (2011), in which several clades were recognized as candidate species, continued with the nuclear data here presented and concluded with the morphological differentiation and characterization of L. caparensis sp. nov. presented here.
Even though there is not a formal description of the morphological characteristics that define the magellanicus group, the features that defined the species Liolaemus magellanicus do differentiate it from the other groups of the L. lineomaculatus section (scale counts and shape, coloration patterns, presence of precloacal pores and size, among others: Scolaro 1992; Laurent 1995; Etheridge & Espinoza 2000; Pincheira-Donoso & Núñez 2005). The divergence of the magellanicus group from the rest of the section was inferred to have occurred during the Late Miocene (~8.46 million years ago, Breitman et al. 2011).
Morphological, molecular, and geographic distributional data provide evidence to consider Liolaemus caparensis sp. nov. a distinct species most closely related to L. magellanicus , forming the magellanicus group. Because the aim of this work was to describe the new species and not to propose new taxonomical arrangements, we avoid hypothesizing new taxonomies or presenting morphological features that might characterize new classifications. Including the species described here and two more species described in another manuscript (Breitman et al. in press), the number of species included in the L. lineomaculatus section increases to 21.
Our research group is studying the Liolaemus lineomaculatus section to delimit species boundaries and reconstruct relationships from multiple sources of evidence. Our goal is to present monographic revisions of poorly known groups on the basis of an “integrative taxonomy” approach ( Padial et al. 2010), based on independent lines of evidence to propose taxonomic re-arrangements and support hypotheses of species limits. In parallel, we are using integrative phylogeographic analyses based on molecular, morphological, ecological, and geographical data, to reconstruct a temporal sequence of demographic histories, and to eventually test for shared patterns of divergence with other Patagonian clades ( Sérsic et al. 2011).
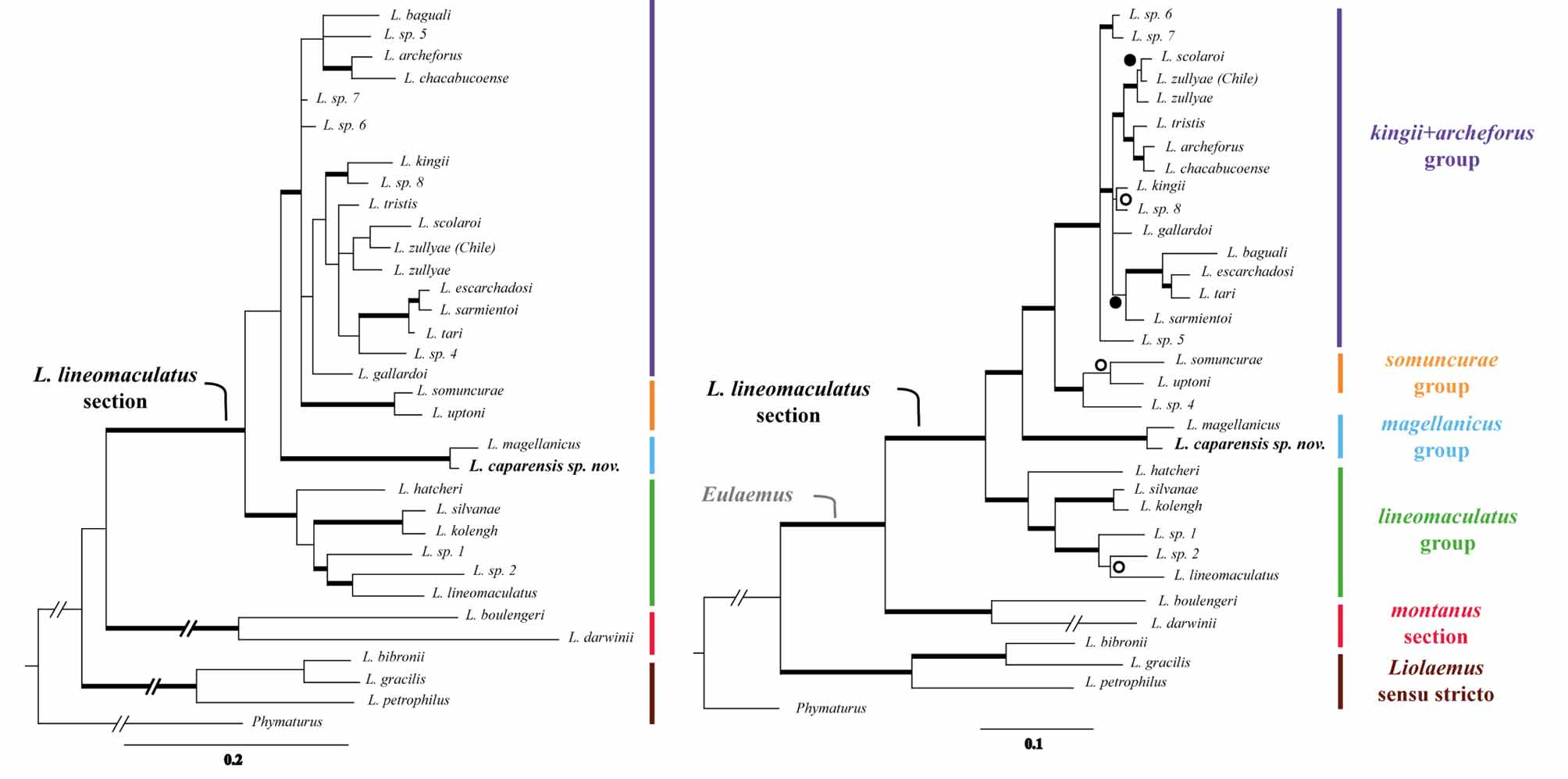


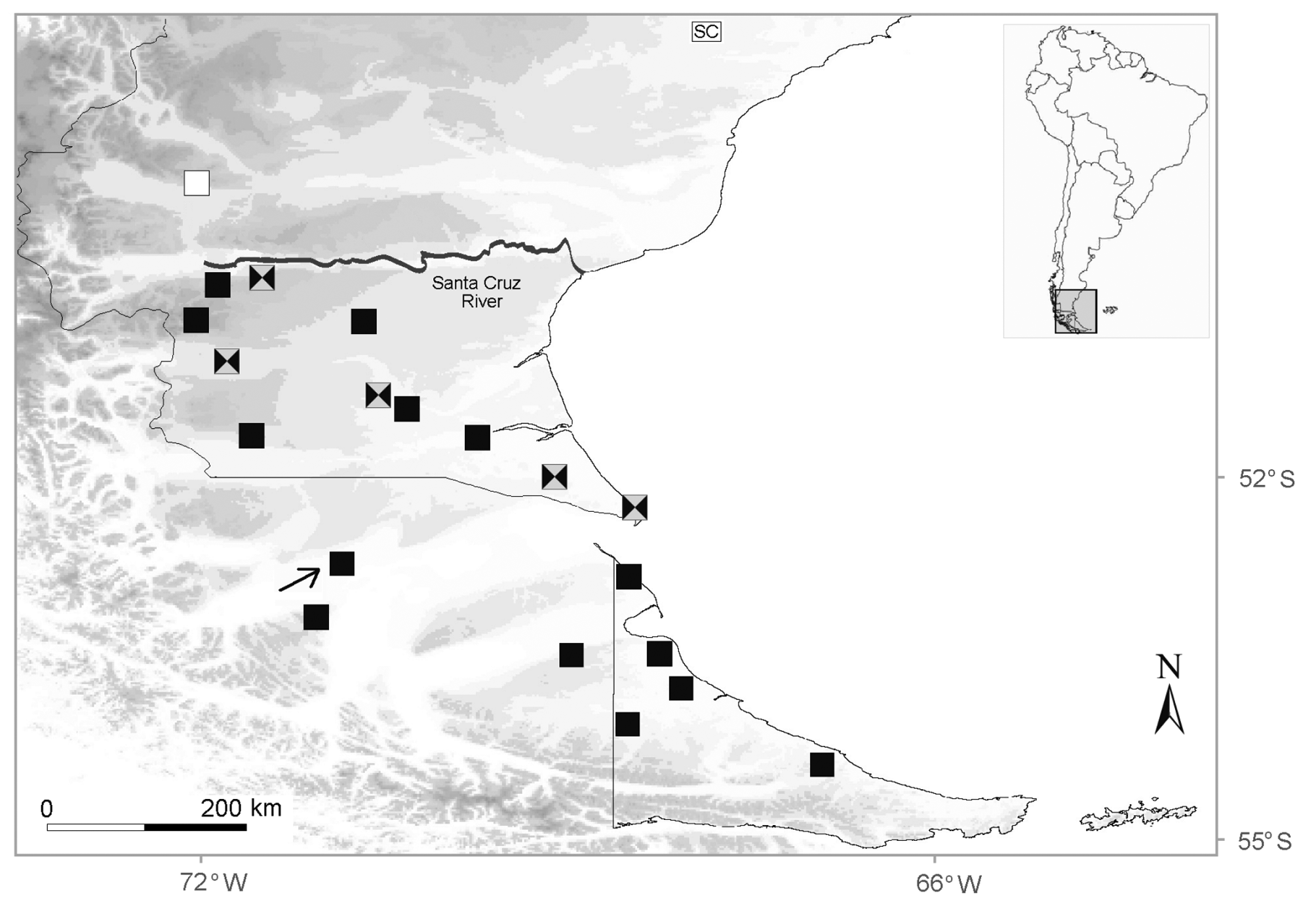
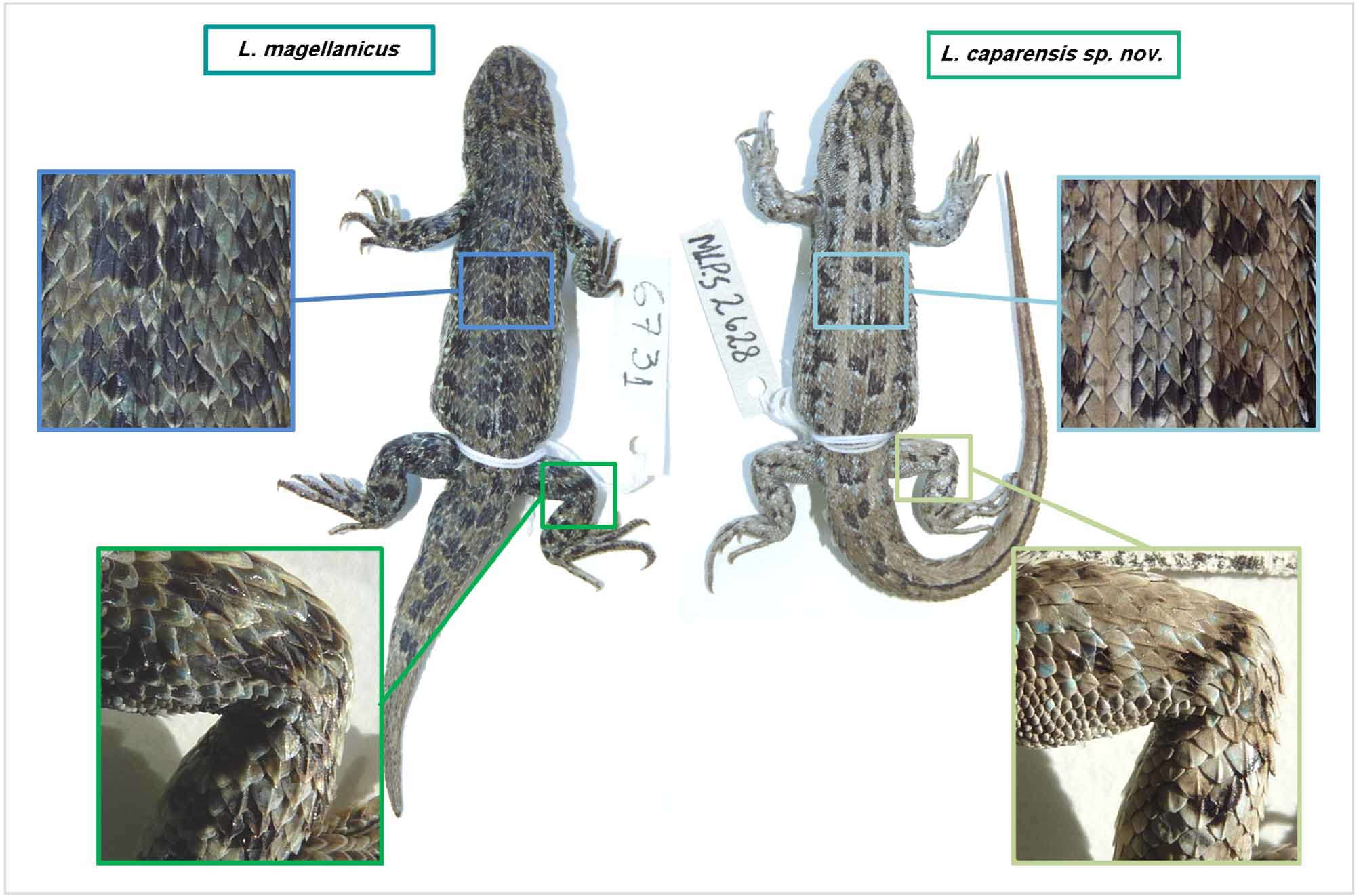
TABLE 5. Variation between females and males of Liolaemus caparensis sp. nov. summarized as mean ± SD (min – max) values for all meristic and morphometric (in millimeters) variables.
| Var | Females (n=5) | Males (n=3) |
|---|---|---|
| SCI | 6.75±1.04 (5–8) | 6±1 (5–7) |
| LS | 5±0.53 (4–6) | 4.33±1.15 (3–5) |
| SS | 6.88±0.83 (6–8) | 6.33±0.58 (6–7) |
| IS | 4.75±0.71 (3–5) | 4.67±0.58 (4–5) |
| MS | 44.25±4.95 (39–55) | 42.67±2.31 (40–44) |
| DS | 42.13±2.59 (39–47) | 39.67±1.15 (39–41) |
| VS | 68.75±3.01 (64–73) | 67.33±3.21 (65–71) |
| IL3 | 14.88±1.36 (14–18) | 15.33±0.58 (15–16) |
| IL4 | 20.25±1.67 (18–23) | 20.5±0.71 (20–21) |
| Pores | 0 | 3.67±0.58 (3–4) |
| SVL | 49.8±2.77 (47–54) | 55±3.61 (52–59) |
| TL | 63±2.94 (60–67) | 74±8.72 (68–84) |
| DFH | 23.84±1.62 (21.2–25.2) | 24.87±0.93 (24.1–25.9) |
| FOL | 12.52±0.71 (11.37–13.19) | 14.99±0.44 (14.68–15.3) |
| TFL | 6.86±0.55 (6.19–7.55) | 7.86±0.72 (7.19–8.62) |
| RUL | 5.08±0.44 (4.58–5.66) | 5.51±0.12 (5.38–5.61) |
| HAL | 7.55±0.53 (7.08–8.36) | 8.53±0.59 (7.85–8.91) |
| HH | 6.16±0.41 (5.65–6.7) | 7.05±0.56 (6.72–7.7) |
| HW | 8.8±0.43 (8.4–9.33) | 10.41±0.91 (9.37–11.07) |
| HL | 10.95±0.37 (10.43–11.4) | 12.38±0.91 (11.69–13.41) |
| EH | 1.79±0.14 (1.58–1.89) | 1.98±0.27 (1.77–2.29) |
| EL | 2.99±0.1 (2.85–3.07) | 3.16±0.24 (2.91–3.38) |
| RND | 1.83±0.22 (1.58–2.15) | 2.16±0.13 (2.01–2.26) |
| RH | 0.97±0.05 (0.9–1.01) | 1±0.1 (0.94–1.12) |
| RL | 2.41±0.16 (2.26–2.66) | 2.83±0.11 (2.71–2.93) |
| DRE | 4.65±0.25 (4.38–4.99) | 5.23±0.22 (5.09–5.49) |
| AH | 1.82±0.18 (1.65–2.09) | 2.13±0.17 (1.94–2.24) |
| AL | 1.58±0.25 (1.33–1.88) | 1.62±0.03 (1.59–1.64) |
| MLP |
Museo de La Plata |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.