
Polypedilum (Tripodura) masudai (Tokunaga)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3893.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:34FE5DA3-C783-4EB6-A2A5-38FB2F5D8DD2 |
|
DOI |
https://doi.org/10.5281/zenodo.3508076 |
|
persistent identifier |
https://treatment.plazi.org/id/0741E555-6D02-FFEB-FF5E-F921FBB8FDD2 |
|
treatment provided by |
Plazi |
|
scientific name |
Polypedilum (Tripodura) masudai (Tokunaga) |
| status |
|
Polypedilum (Tripodura) masudai (Tokunaga) View in CoL
Chironomus (Polypedilum) masudai Tokunaga, 1938: 331 View in CoL .
Polypedilum masudai Tokunaga View in CoL : Sasa, 1985: 44; Sasa & Kikuchi, 1986: 24; Ree & Kim 1988: 14. Polypedilum (Tripodura) masudai Tokunaga View in CoL : Sasa, 1991: 84.
Material examined. 1P♂ and 2Le, Macau, the Bay of St. Lazarus, near Taipa Houses Museum, 14. xii. 2013 (H.Q. Tang); 2Pe and 8L, same locality, 27. ii. 2013, (H.Q. Tang); 35Pe, Macau, artificial ecology park of Alto de Coloane, 18. i. 2013 (H.Q. Tang); 3P♂ and 2L, China, Guangxi, Hezhou city, Fulong, 10. viii. 2013 (J.G. Zhao) (all EJNU).
Larva (n=4). Head capsule yellow, with dark-brown postmentum. Mentum and mandible teeth dark brown. Occipital margin dark.
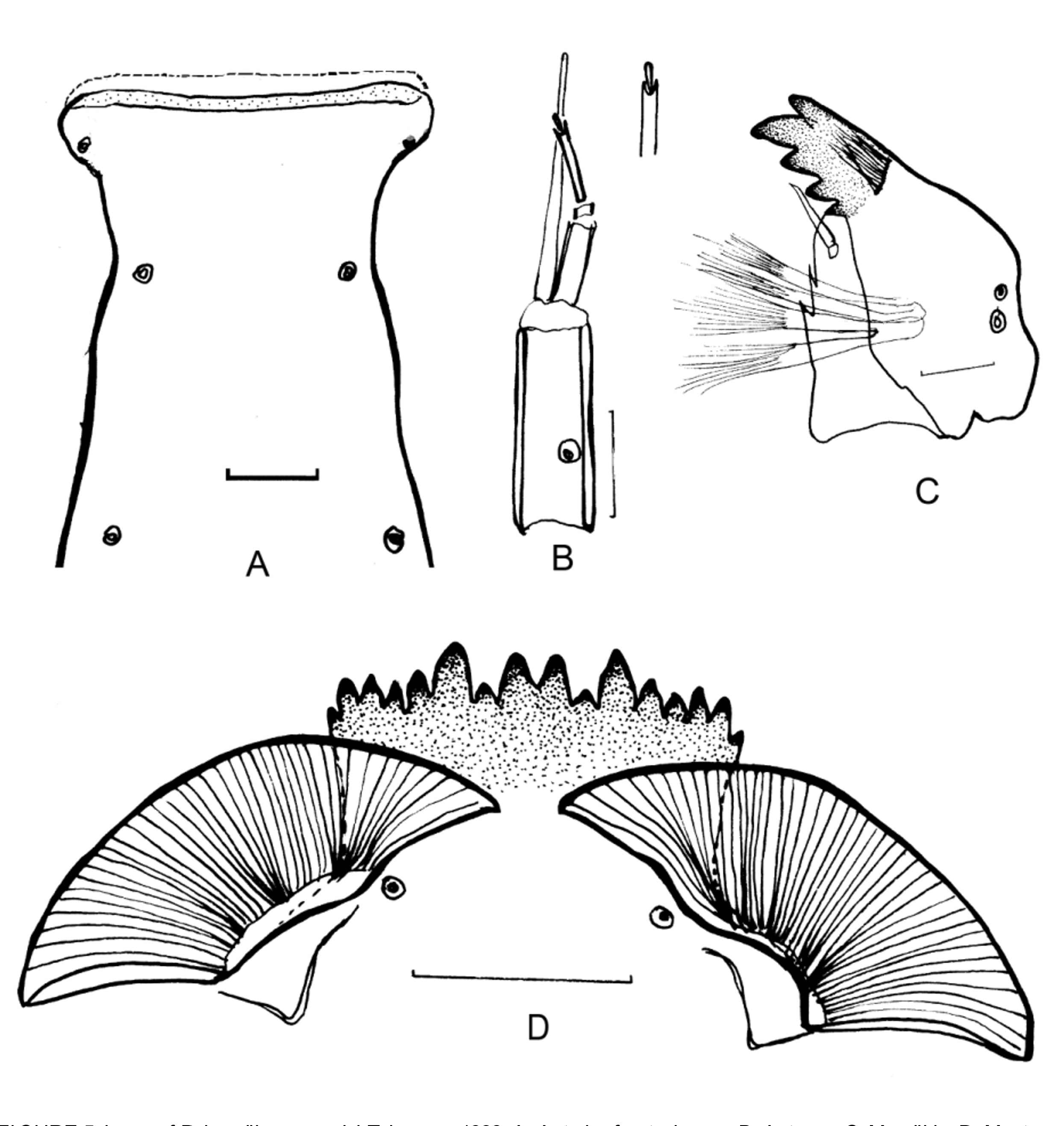
Dorsal surface of head ( Fig. 5 View FIGURE 5 A). Frontoclypeus with a narrow hyaline band, surrounded the broadened apex.
Antenna ( Fig. 5 View FIGURE 5 B) pale yellow, with reduced third and fifth segment, segment 2 and 4 relative long. Antennal blade extending beyond the terminal of flagellum.
Labrum. SI and SII fine plumose. Pecten epipharyngis consists of 3 scales, each scale with 3 teeth. Premandible with 2 apical teeth and brush present.
Mandible ( Fig. 5 View FIGURE 5 C). With 1 apical, 1 dorsal and 2 inner teeth, the length of apical tooth subequal to the combined width of 2 inner teeth. Seta subdentalis almost reaching the second inner teeth. Mola with 2–3 sharp spines.
Mentum ( Fig. 5 View FIGURE 5 D) with 2 median teeth and 7 laterals, the location of outermost laterals relative low, the fourth laterals lower than the neighboring teeth giving the appearance of the relative greater elevation of fifth and sixth laterals. Ventromentum wider than the mentum, with the medially-directed pointed apex. Striae crowded in dense basally, 5–6 striae about 5 µm wide, total about 40 striae. Inter-plate distance about subequal to the width of two median teeth.
Body. Procercus golden brown, with 7–8 anal seta. Anal tube with a constriction in the middle. Mensural features as in Table 1 View TABLE 1 .
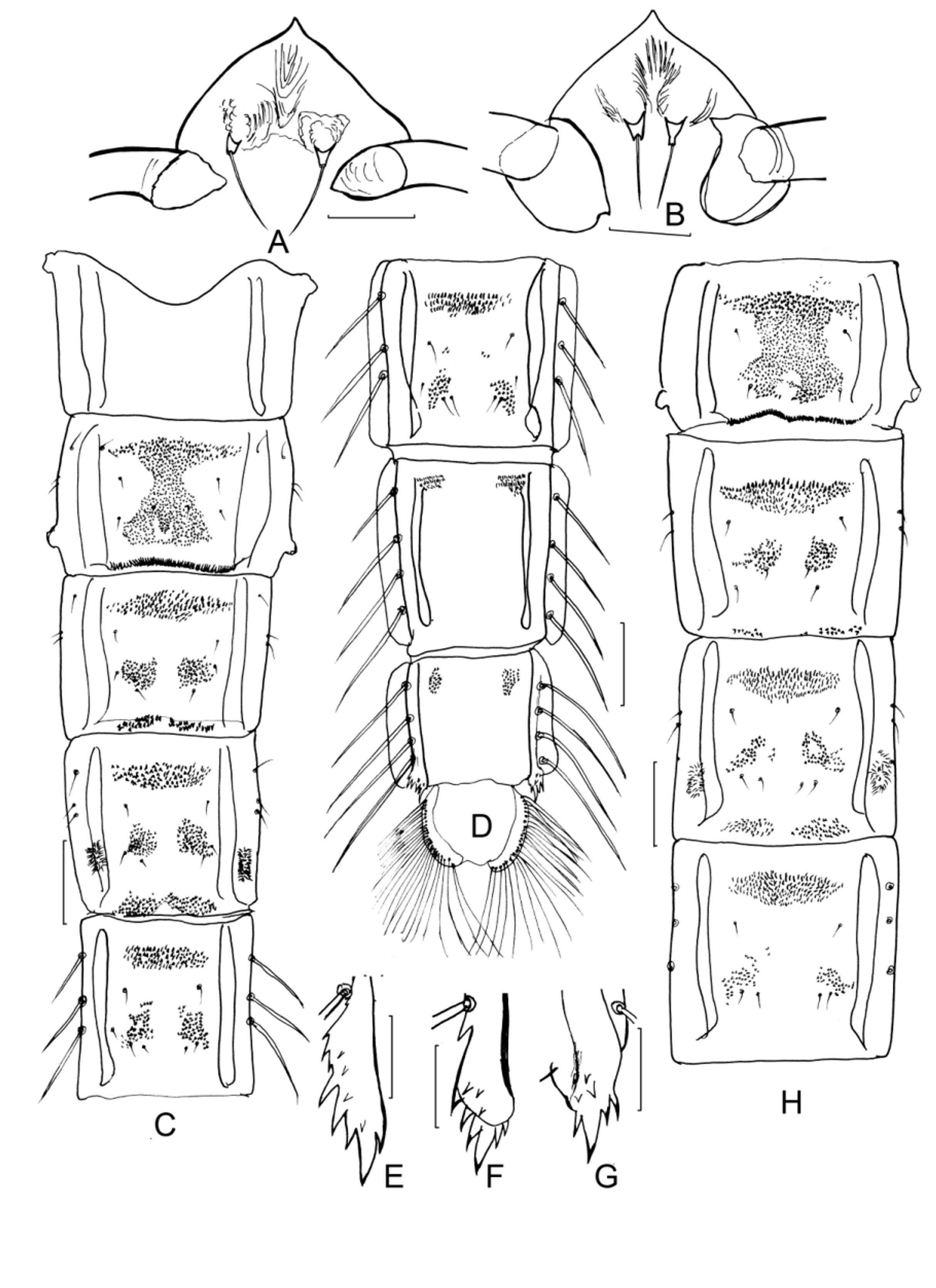
Pupa (n=5). Pupal exuviae pale yellow, apophyses brown. Cephalic tubercles present, the height equal to the width; frontal setae long, more than 2 times as long as the height of cephalic tubercles. Mid-thorax with one row of sparse granules near suture.
Abdomen ( Fig. 6 View FIGURE 6 C & 6D). Hook row II 0.47–0.63, 0.56 times as wide as corresponding tergite width. Tergites II–VI with obvious anterior transverse bands of points stronger than those of the posterior patch and conjunctive patch, median armament greatly reduced. Posterior patch of tergites III–VI consist of 2 separated sub-patches. Spinulation of tergite II continuous, a relative larger anterior transverse band of points fused broadly with the postero-median trapezoidal patch of small spinules. Tergite VII and VIII only with 2 anterolateral small patches, without trace of median patch.
Conjunctives present in tergite III/IV and IV/V, with spinules about the same size as that of median bands, medially usually interrupted in conjunctives III/IV or weakly connected in IV/V. The number of teeth in conjunctive III/IV varied greatly, ranging from 9–36 and 8–30 in two parts separately, the gap about half width to whole width of each part ( Fig. 6 View FIGURE 6 C & H). Comb of segment VIII composed of 3–8 golden spines ( Fig. 6 View FIGURE 6 E–G), often with one largest spine posteriorly. Lateral taeniae of segments IV–VIII: 0, 3, 3, 4, 4. Mensural features as in Table 2 View TABLE 2 .
Remarks. Larvae of P. masudai cannot be separated with certainty from other members of P. bicrenatum group in Europe (Kiknidze et al. 1999, Klink 2002) or P. halterale group in North America ( Epler 2001) which has a similar reduced third and fifth antennal segment. However, the heavy sclerotized dark postmentum and the relative low position of 4th mental laterals is distinctive compared to all described Tripodura larvae. Pupae of P. masudai resembles P. digitifer Townes 1945 in the reduced median spinulation on tergites III–VI and posterior transverse band with the relatively well separated two patches, but the latter can be separated by the reduced cephalic tubercles ( Soponis & Simpson 1992).
Biology. Larvae of P. nodosum are found in the bottom sediment of lentic waterbodies ( Johannsen 1932, Tokunaga 1964, Sasa & Hasegawa 1983). Our collections from the Macau ponds came from substrates consisting of dense detritus and fine mud, perhaps representing a mesotrophic to eutrophic condition. Other common chironomids in these ponds included Chironomus flaviplumus Tokunaga 1940 , Glyptotendipes tokunagai Sasa 1979 , Harnischia longispuria Wang & Zheng 1993 , Tanytarsus formosanus Kieffer 1912 and T. oscillans Johannsen 1932 . Larvae occur massively in shallow areas during colder months, but in deeper areas during the summer.
In Thailand a survey across hundreds of water bodies, including standing, running and waterfalls, larval P. nodosum were found only in Andaman Sea coastal pools that were created as post-impact features of the 2004 Indian Ocean tsunami ( Cranston 2007, Sites & Vitheepradit 2010). Sites varied in conductivity from 45 to 1650 µS and with dissolved solids from 23 to 821 ppm. The species was not found in any unimpacted site across the whole country ( Cranston 2007). Evidently the species tolerates high levels of salinity and nutrients, but also can be a pioneering or colonizing species, being found in artificial tanks and water storage reservoirs in Singapore and artificial research channels in Queensland, Australia (P.S. Cranston pers. obs.). The species is found only in warm waters ( Thailand 28–32°C) and is associated generally with tropical to subtropical regions of Asia and Australasia.
After checking all the published collecting sites from Okinawa to Queensland, we found that the distribution of P. nodosum is almost entirely restricted to the lentic habitat where the site is very close to ocean or coastal regions. All collected dates of pupae and adults indicated that the species is multivoltine, emerging from autumn to the next spring.
Specimens of P. masudai described here were collected from Macau ponds and the Hejiang River, Guangxi, thus it perhaps can inhabit both lentic and lotic water bodies. Those sites usually had muddy to mud-sandy substrates. Based on the seasonal investigation in Macau, it appears that P. masudai is one of the relatively abundant macroinvertebrate species in the cold months. Emergence records indicate that the species is multivoltine. Adults emerge from January to March in Macau and in August in Hejiang, Guangxi.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Chironominae |
|
Genus |
Polypedilum (Tripodura) masudai (Tokunaga)
| Tang, Hong-Qu, Cranston, Peter S., Zhao, Jian-Gang, Lok, Chan-Wa, Wong, Kai-Chin & Li, Zhi-Qiang 2014 |
Polypedilum masudai
| Sasa 1991: 84 |
| Ree 1988: 14 |
| Sasa 1986: 24 |
| Sasa 1985: 44 |
Chironomus (Polypedilum) masudai
| Tokunaga 1938: 331 |